今天是:

目录
插图清单
图3-7 2020-2021年西凉湖各季节不同湖区TLI指数 29
图3-15 2017-2020年西凉湖水质分析PCA图 37
图3-16 2017-2019年西凉湖浮游植物种类组成比例 41
图3-17 2017-2019西凉湖浮游植物密度与生物量 42
图3-18 2017-2019年西凉湖浮游植物密度组成百分比 43
图3-19 2017-2019年西凉湖浮游植物密度组成 44
图3-20 2017-2019西凉湖浮游植物生物量组成百分比 44
图3-21 2017-2019西凉湖浮游植物生物量组成百分比 45
图3-22 2017-2019年西凉湖浮游动物种类组成比例 49
图3-23 2017-2019西凉湖浮游动物密度与生物量 50
图3-24 2017-2019年西凉湖浮游动物密度组成 52
图3-25 2017-2019年西凉湖浮游动物密度组成百分比 52
图3-26 2017-2019年西凉湖浮游动物生物量百分比 53
图3-27 2017-2019年西凉湖浮游动物生物量组成 53
图3-28 2017-2019年西凉湖底栖动物种类组成百分比 56
图3-29 2017-2019年西凉湖底栖生物密度与生物量 57
图3-30 2017-2019年西凉湖底栖动物密度组成 59
图3-31 2017-2019年西凉湖底栖动物密度百分比 59
图3-32 2017-2019年西凉湖底栖动物生物量组成 60
图3-33 2017-2019年西凉湖底栖动物生物量百分比 60
图3-40 2020-2021年西凉湖鱼类活动水层组成(密度) 69
图3-41 2020-2021年西凉湖鱼类活动水层分布(生物量) 69
图3-42 2020-2021年西凉湖鱼类食性组成(密度) 70
图3-43 2020-2021年西凉湖鱼类食性组成(生物量) 71
图3-44 2020-2021年西凉湖鱼类繁殖习性组成(密度) 72
图3-45 2020-2021年西凉湖鱼类繁殖习性组成(生物量) 72
图3-46 2020-2021年西凉湖鱼类受精卵类型组成(密度) 73
图3-47 2020-2021年西凉湖鱼类受精卵类型组成(生物量) 73
图3-48 2020-2021年西凉湖鱼类群落PCoA分析 76
附表清单
表3-4 2020-2021年西凉湖各水域水体理化特征季节分布 20
表3-5 中国湖泊(水库)部分参数与Chl-a的相关关系rij及rij2值 27
表3-7 2020-2021年西凉湖各季节不同湖区营养状况综合评价 28
表3-9 2017-2020年秋季西凉湖水体富营养程度 35
表3-11 2017-2019年西凉湖浮游植物平均密度组成 42
表3-12 2017-2019年西凉湖浮游植物生物量组成 42
表3-15 2017-2019年西凉湖浮游动物平均密度组成 50
表3-16 2017-2019年西凉湖浮游动物生物量组成 51
表3-19 2017-2019年西凉湖底栖动物密度组成 57
表3-20 2017-2019年西凉湖底栖动物生物量组成 58
表3-29 2017-2019年西凉湖浮游生物鱼产力估算 79
表3-30 2017-2019年西凉湖底栖生物鱼产力估算 79
表3-31 2017-2019年西凉湖高等水生植物鱼产力估算 80
表4-13 2021-2025年西凉湖鱼类增殖放流清单 130
西凉湖位于湖北省咸宁市西北方(E114°00′-114°10′,N29°51′-30°02′),属长江中游浅水草型湖泊,湖区面积12.78万亩,为湖北省第五大湖泊,是咸宁市第一大湖泊,呈裤状型,跨咸安、嘉鱼、赤壁三县市区,沿湖周边有向阳湖、渡普、新街、官桥、神山、官塘驿六个乡镇,15个村,湖区人口达3.8万人。分东西凉湖与西西凉湖,东西凉湖东、东北、南面丘陵,西北面平原,西西凉湖东、南、北面丘岗,西面平原,西西凉湖略高于东西凉湖。沿湖还有若干小湖汊,西港湖、思姑台,伯良湖、马狮湖、泉口湖、钟鸣湖,在20世纪70年代,嘉鱼和赤壁分别将以上湖汊由人工筑堤隔断,(伯良湖2020年已退垸还湖拆掉一部分石头堤)面积达到将近2万亩,使西凉湖的面积不断减少。
东西凉湖源头有汀泗河、泉口河,西西凉湖有宋家河,三条河河水流入西凉湖,汇聚到嘉鱼渡普望东庙前面的竞江口出湖口注入余码河流入长江,余码头建有通江大闸,可调控西凉湖水位,另外也可通过金水河在江夏的金口文昌阁注入长江。西凉湖湖水平均2.1 m,最大深度达8.0 m,湖长19.1 km,平均宽度3.5 km,最大宽度8.8 km,沿湖长377.0 km,设防水位21.5 m,警戒水位23.0 m,保证水位24.0 m。 西凉湖具有调蓄、灌溉、泄洪、气候调节、生活供水、旅游航运、物种保护等多种功能,是长江中下游极具典型性的天然生物资源多样性湖泊,在长江水域生态网中具有十分重要的地位。由于西凉湖湖面壮阔,湖汊众多,水生物丰富,千百年来,一直是鱼鸟栖息的天堂,中国大地上珍贵的湿地。
西凉湖流域属于鄂东南低山丘陵区向江汉平原东部边缘过渡。地势是南高北低,岭谷平行相间,山丘盆地参差,沿江滨湖交接。西凉湖南部边缘属幕阜山脉,自西南向东北延绵,海拔1000米左右。流域水系发育良好,水量丰富,以成河谷沉溺湖类为主,横断面呈开阔的“U”字形。
湖区属于典型的亚热带大陆性季风气候,夏热冬冷,四季分明,雨量充沛,气候温和。由于幕阜山脉面对季风温暖气流来向和地形抬高作用,促成西凉湖流域成为湖北省的多雨和暴雨中心。流域多年平均降水量为1300-1390毫米。其中,夏秋降雨量大,特别是梅雨期多降暴雨、大暴雨,汛期(5-9月)的降雨了量在700毫米左右,占全年的50%左右。最大暴雨量一般出现在6月中下旬-7月中上旬,但丰水年、枯水年之间差别大。境内多年平均气温16.8-16.9°C,极端最高气温39.7°C,极端最低气温-12°C,年无霜期259-261天,平均日照时数1524-1945小时。
历史上西凉湖水清澈见底,水草茂盛,水体基本无污染,但在1980年以后,随着乡镇企业发展和村镇人口增多,大量未经处理的污水直接排入湖内,污染程度较大。21世纪之前,湖区内过度“围湖造田、围网养殖”等破坏水生态环境的行为,造成湖区内污染严重。2020年经西凉湖排口调查共有77个,其中涵闸类排口14个,泵站类排口2个,明渠类排口33个,管道类排口23个,其他类排口5个。
2000年以前,湖区虽有管理机构,但尽管渔业生产,环保事项基本搁置,但过度“围湖造田、围网养殖”等破坏水生态环境的行为,造成天然湖域减少,天然水面减少,天然水生物资源减少。2001年以来,西凉湖环保工作提上议事日程。2008年7月,农业部批复建立西凉湖国家鳜鱼、黄颡鱼水产种质自然保护区。2009年12月,咸宁市政府召开专题会议,研究建立西凉湖市级水生生物自然保护区问题。2010年8月,咸宁市人民政府正式批准西凉湖为市级水生生物自然保护区。2012年,西凉湖管理局正式启动申报西凉湖省级自然保护区事宜,省、市政府将湖区建设纳入省级和地方建设总体规划之中,通过合理规划,理顺管理体制,实行统一管理,有计划地引导湖区人民调整不合理产业模式。2011 年,咸宁市启动西凉湖水生态修复工作,出台西凉湖水生态修复方案(2011-2015年),并投资850万元,开展灌江纳苗和人工增殖放流,补种沉水、挺水、浮叶植物,投放贝类等两栖生物。从2010 年起,沿湖 6 个乡镇加强污水和垃圾处理工作,加紧建设地埋式无动力污水处理装置,同时加紧建设集中式城镇污水处理厂,强化工矿企业排污监管,限期停产、转产污染较重企业,或搬迁到县、市、区工业园区。2010 年,赤壁市与咸安区联合拆除汀泗河上游双石水库2公顷网箱养鱼,避免了投饵养殖污染水体。种种措施使西凉湖水质、水生态环境得到改善,遏制西凉湖水域水质恶化趋势。2017年,完成了9.3万亩的水面围网拆除任务,破碎化的湖面得以恢复,生态环境得到极大的改善。然而,目前湖区内鱼类资源仍处于匮乏状态,鱼类群落多样性较低,沉水植物种类以及数量仍处在恢复期,生态系统完整性较差,物质和能量转换效率较低。
为全面贯彻落实国务院《水污染防治行动计划》(国发[2015]17 号)和湖北省《水污染防治行动计划工作方案》(鄂政发 [2016]3 号),为完成水污染防治工作目标任务及重点流域水污染防治“十三五”规划目标,根据西凉湖所在区域、公众影响和污染程度,需开展水体生态系统结构与功能修复,改善水环境质量。因此,本规划贯彻了水污染防治行动计划工作方案的精神,大力推进了生态文明的建设。
《渔业法》和《中国水生生物资源养护行动纲要》对水生生物资源养护提出了明确要求,开展生物资源养护是保护水生物资源的重要途径之一,坚持在利用中保护、在保护中利用的发展思路,最终实现水域生态和渔业生产相互协调、生态保护和渔业发展相得益彰。本规划着眼于水生生物资源养护,集成了协同的水生态修复技术体系,保障了水生生物资源恢复效果。
大水面渔业是湖北省渔业的重要组成,为水域生态文明建设、保障优质水产品供给、推进产业融合、促进农渔民增收作出了重要贡献,但发展方式亟待转型升级。《农业农村部 生态环境部 国家林业和草原局关于推进大水面生态渔业发展的指导意见》(农渔发〔2019〕28号),以下简称《意见》)明确了大水面生态渔业发展目标,聚焦解决当前大水面生态渔业发展中存在的突出矛盾和问题,提出了具有针对性和可操作性的具体措施,是当前和今后一个时期推进大水面生态渔业发展的布局依据。本规划贯彻了《意见》精神,满足了发展大水面生态渔业的需求。
修复城市湖泊生态景观,形成以湖泊景观为核心,以湖泊周边湿地及绿地为屏障,以外围的山林、城市公园、大型广场绿地、环城林带等形式的绿地系统为基础,以连接湖泊的各种水渠为生态走廊,突出以湖为中心,以河为纽带,以水为媒介的城市旅游资源,形成一种善水的旅游氛围,使城市人文景观和自然景观有机融合,提升城市湖泊的观赏价值,必然会增强城市旅游业的开发实力,刺激城市旅游业的发展,为咸宁市的经济发展开辟新的亮点。
以习近平生态文明思想为指导,以改善湖泊生态环境质量为核心,编制《咸宁市西凉湖水生态修复与保护发展规划(2021-2025)》,科学指导“十四五”期间西凉湖生态修复与保护工作,持续加大西凉湖生态修复力度,加强水生生物多样性保护。在保护中利用,在利用中保护,开展“绿水青山就是金山银山”创新实践,协同推进地区经济高质量发展。
(1)《中华人民共和国渔业法》(2013年12月28日修订)
(2)《中华人民共和国水法》(2016年7月2日修订)
(3)《中华人民共和国水污染防治法》(2008年2月28日修订)
(4)《中华人民共和国环境影响评价法》(2016年7月2日修订)
(5)《中华人民共和国水产资源繁殖保护条例》(1979年2月10日起颁布施行)
(6)《中华人民共和国水生动植物自然保护区管理办法》(2014年4月25日修订)
(7)《中华人民共和国水生野生动物利用特许办法》(1999年9月1日起施行)
(8)《长江渔业资源管理规定》(1995年9月28日起施行)
(9)《水产苗种管理办法》(2005年4月1日起施行)
(10)《渔业水域污染事故调查处理程序规定》(1997年3月26日起施行)
(11)《湖北省湖泊保护条例》(2012年5月30日颁布施行)
(12)《湖北省水污染防治条例》(2014年1月22日颁布施行)
(1)《中共中央国务院关于加快推进生态文明建设的意见》(2015年4月25日,中发〔2015〕12号)
(2)《国务院办公厅关于加强长江水生生物保护工作的意见》(2018年10月15日,国办发〔2018〕95号)
(3)《国务院关于印发水污染防治行动计划的通知》(2015年4月2日,国发〔2015〕17号)
(4)《全国生态环境保护纲要》(2000年11月26日,国发〔2000〕38号)
(5)《农业农村部等10部委关于加快推进水产养殖业绿色发展的若干意见》(2019年2月15日,农渔发[2019]1号)
(6)《水产种质资源保护区的管理暂行办法》(2011年3月1日,农业部令2011年第1号)
(7)《农业部关于推动落实长江流域水生生物保护区全面禁捕工作的意见》(农长渔发〔2017〕1号)
(8)《农业农村部 生态环境部 国家林业和草原局关于推进大水面生态渔业发展的指导意见》(农渔发〔2019〕28号)
(9)《中国水生生物资源养护行动纲要》(国发 〔2006〕9号)
SC/T 1149-2020 大水面增养殖容量计算方法
SL 219-2013 水环境监测规范
GB 13195-91 水质 水温的测定-温度计或颠倒温度计测定法
SL 87-1994 透明度的测定
GB 11893-1989 水质 总磷的测定 钼酸铵分光光度法
GB 11894-1989 水质 总氮的测定 碱性过硫酸钾消解紫外分光光度法
SL 88-2012 水质 叶绿素的测定 分光光度法
GB 11892-89 水质 高锰酸盐指数的测定
DB42/T 1174-2016 湖泊鳜放养技术规范
DB42/T 1177-2016 湖泊鳙放养技术规范
DB42/T 1178-2016 湖泊蒙古鲌放养技术规范
西凉湖,面积85.20 km2。
规划期限为5年,即2021-2025年。
规划编制组于2017年12月、2018年11月、2019年11月、2020年11月连续四年对西凉湖水质间年际变化的监测同时分析了2017-2019年浮游生物、底栖生物等方面;2020-2021年的水环境与渔业资源进行了同步监测,分析了水质、水生植物、鱼类等方面,掌握了西凉湖生态环境演变特征与趋势,查明了其存在的生态环境问题,为下一步生态修复方案编制提供了数据支撑和问题导向。
3.1.1.1 水质指标
水质指标包括水深、透明度、浊度、水温、pH、电导率、溶解氧、总磷、总氮、高锰酸盐指数、氨氮等。
3.1.1.2 水生生物
浮游植物的种类组成、密度与生物量、优势种种类及生物量等;浮游动物的种类组成、密度与生物量、优势种种类及生物量等;底栖动物的种类组成、密度与生物量、优势种种类及生物量等;水生高等植物包括种类组成、密度、优势种种类以及空间分布情况;鱼类的种类组成、密度与生物量、优势种种类及生物量等。
2017-2019年在西凉湖西湖区共设置6个采样点(图3-1,XLH1-6),2020-2021年全湖共设置15个采样点(图3-1,XLH1-15),其中东湖区6个采样点,西湖区6个采样点,中间水道3个采样点,各点坐标见表3-1。2017-2020年每个采样点监测水质,2017-2019年监测浮游植物、浮游动物和底栖动物等生物指标,2020-2021年每个采样点对水质鱼类群落以及水生植物进行调查,各点坐标见表3-1。
样点 | 坐标 | 样点 | 坐标 |
XLH1 | 114°2′27.78"E 29°54′46.05"N | XLH9 | 114°4′38.11"E 29°58′42.59"N |
XLH2 | 114°2′28.50"E 29°55′55.86"N | XLH10 | 114°5′38.05"E 29°58′46.97"N |
XLH3 | 114°3′42.89"E 29°56′10.26"N | XLH11 | 114°6′59.25"E 29°58′36.86"N |
XLH4 | 114°3′4.88"E 29°56′48.04"N | XLH12 | 114°6′2.58"E 29°58′3.10"N |
XLH5 | 114°4′2.94"E 29°57′11.24"N | XLH13 | 114°6′45.94"E 29°57′34.33"N |
XLH6 | 114°3′3.48"E 29°57′47.49"N | XLH14 | 114°7′9.66"E 29°56′36.5N |
XLH7 | 114°3′47.04"E 29°58′9.54"N | XLH15 | 114°7′42.61"E 29°55′49.87"N |
XLH8 | 114°4′2.82"E 29°58′47.12"N |

参照《渔业生态环境监测技术规范-淡水》(SC/T 9102.3-2007)、《内陆水域渔业自然资源调查手册》、《水环境监测规范》(SL 219-2013)、《地表水环境质量评价办法》(GB3838-2002)等技术资料描述的方法进行调查监测。
3.1.3.1 水质
现场测定水深、透明度、水温、浊度、电导率、pH以及溶解氧等指标;各样点水样用干净的1 L塑料瓶密封避光保存,立即带回实验室测定总磷(TP)、总氮(TN)、叶绿素a、高锰酸盐指数(COD)、氨氮(NH4+)、亚硝态氮(NO2-)、硝态氮(NO3-)、正磷酸盐(PO43-)含量。具体方法见表3-2。
参数 | 采样方法 | 测试方法 | 方法来源 |
水温 | 现场测量 | 现场测定 | YSI Pro DSS 多参数水质测定分析仪 |
pH | 现场测量 | 现场测定 | YSI Pro DSS 多参数水质测定分析仪 |
溶解氧 | 现场测量 | 现场测定 | YSI Pro DSS 多参数水质测定分析仪 |
浊度 | 现场测量 | 现场测定 | YSI Pro DSS 多参数水质测定分析仪 |
透明度 | 现场测量 | 塞氏盘法 | 《透明度的测定》(SL-87-1994) |
水深 | 现场测量 | 现场测定 | SM-5便携式测深仪 |
高锰酸盐数 | 用相应容器采集足量水样后加入相应固定剂进行固定,低温保存运输回实验室进行下一步分析 | 滴定法 | 水质 高锰酸盐指数的测定(GB 11892-89) |
总磷 | 钼酸铵分光光度法 | 水质 总磷的测定 钼酸铵分光光度法(GB 11893-1989) | |
总氮 | 碱性过硫酸钾消解紫外分光光度法 | 水质 总氮的测定 碱性过硫酸钾消解紫外分光光度法(GB 11894-1989) | |
氨氮 | 纳氏试剂分光光度法 | 水质 氨氮的测定 纳氏试剂分光光度法(HJ 535-2009) | |
硝态氮 | 紫外分光光度法 | 水质 硝酸盐氮的测定 紫外分光光度法(HJ-T 346-2007) | |
亚硝态氮 | 分光光度法 | 水质 亚硝酸盐氮的测定 分光光度法(GB/T 7493-1987) | |
正磷酸盐 | 分光光度法 | 生活饮用水标准检验方法 无机非金属指标(GB/T 5750.5-2006) | |
叶绿素a | 分光光度法 | 水质叶绿素的测定分光光度法(SL88-2012) |
3.1.3.2 浮游植物
取样时,定性样品用25号浮游生物网在水面下0-0.5 m处作“∞”形循回拖动,3-5 min后将网慢慢提起,将浓缩的水样放入标本瓶中,取样约30-50 mL,用1% -1.5%样品体积的鲁哥试液固定。定量样品用采水器在所测水层采水1000-2000 mL,每升加入10-15 mL鲁哥氏液充分摇匀固定,实验室内静止48小时后浓缩至30 mL。
镜检时,取0.1 mL样品,放入0.1 mL计数框内,盖上盖玻片。在200-400倍镜下,利用视野计数法或全片计数法进行定量检测。在进行镜检计数时,每个样品不低于2个片子,当两片的计数值时,取其平均值作为测定结果。当两片误差超过15%时,进行第三片计数,取结果相近的两片(误差小于15%)的平均值作为测定结果。
3.1.3.3 浮游动物
定量样品用5 L的采水器分次取20 L水样,再用25号浮游生物网过滤浓缩,过滤物放入50 ml标本瓶中,并用滤出水洗过滤网3次,所得过滤物也放入50 ml标本瓶中,此为枝角类和桡足类的定量样品;原生动物、轮虫和无节幼体定量用浮游植物的定量样品。定性样品用13号浮游生物网在表层缓慢拖曳采集。注意定量和定性样品的采集网要分开使用。在标本瓶中加入5%的甲醛溶液约5 ml杀死并固定浮游动物。
镜检时,原生动物、轮虫和无节幼体的鉴定即利用浮游植物样品,吸出0.1 mL样品,置于0.1 mL计数框内,盖上盖玻片,在中倍显微镜下全片计数。每瓶样品计数两片,取平均值;对于轮虫:利用浮游植物样品,吸出1 mL样品,置于1 mL计数框内,全片计数。每瓶样品计数两片,取平均值;对于枝角类、桡足类,用5 mL计数框将样品分若干次全部计数。如样品中个体数量太多,将样品稀释至30 mL或50 mL,用5 mL计数框全片计数。每瓶样品计数两片。
3.1.3.4 底栖动物
用1/16 m2的彼得森采泥器进行定量采样,泥样经16目/cm2的筛网筛洗后在解剖盘中将底栖动物活体标本检出,用10%的福尔马林固定后在实验室中进行镜检鉴定、计数和称重,并换算成单位面积(m2)底栖动物的密度和生物量。
3.1.3.5 高等水生植物
使用1/8 m2抓斗式采草器进行定量采样,水生植物经采取并清洗干净后,对其种类进行鉴别,并对其重量进行记录。相关鉴定依据包括《中国水生植物》和《水生植物图鉴》等
3.1.3.6 鱼类资源
对于鱼类区系组成,采用渔获物调查、文献资料收集等。将渔获物调查的鱼类标本进行现场鉴定或保存后待会室内鉴定,鉴定依据包括《中国动物志 硬骨鱼纲》和《长江鱼类》等。
在西凉湖共设置15个渔获物采样点,东湖区6个、西湖区6个以及水道3个,分别在2020年11月(秋季)、2021年4月(春季)通过定制多网目复合刺网和地笼进行定量采集渔获物,分析种类组成、相对丰度、生物量参数等,采集鳞片等年龄鉴定材料,并记录鱼类体长、体重等信息。其中多网目复合刺网主要是参考Appelberg(2000)所提出的方法,共分为12个网目,高2.0 m,长30.0 m,各网目大小分别为1.0 cm、1.6 cm、2.0 cm、2.5 cm、3.1 cm、4.0 cm、4.8 cm、6.0 cm、7.5 cm、8.5 cm、11.0 cm、12.5 cm;地笼的规格为长15 m,宽0.4 m,高0.3 m,网眼大小为0.5 cm,两侧各有20个入口。
3.1.3.7 多样性指数
运用Shannon-wiener多样性指数、Simpson优势度指数、Pielous均匀度指数、Margalef种类丰富度指数等指标对浮游植物、浮游动物、底栖动物和鱼类群落的物种多样性进行定量分析。具体计算公式如下所:
1)Shannon-wiener多样性指数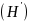
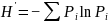
2)Simpson优势度指数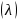
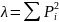
3)Pielous均匀度指数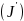
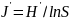
4)Margalef种类丰富度指数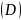
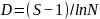
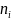 为群落中第i种的个体数,
为群落中第i种的个体数,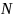 为群落中所有物种数量;
为群落中所有物种数量;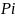 为群落中第i种个体数的比例,即
为群落中第i种个体数的比例,即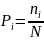 ,
,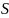 为群落中第i种个体数的比例。
为群落中第i种个体数的比例。
2020-2021年,水体理化特征如表3-3、3-4所示。
3.2.1.1 水深
2020-2021年,全湖平均水深为2.0 m。其中,秋季西湖区平均水深3.3 m、水道平均水深2.6 m、东湖区平均水深3.1 m;冬季西湖区平均水深1.5 m、水道平均水深0.6 m、东湖区平均水深0.8 m;春季西湖区平均水深2.5 m、水道平均水深1.8 m、东湖区平均水深2.1 m,总体上呈现西湖区平均水深高于东湖区。
3.2.1.2 透明度
2020-2021年,全湖平均透明度为67.3 cm。其中,秋季西湖区平均透明度54.2 cm、水道平均透明度53.3 cm、东湖区平均透明度59.2 cm;冬季西湖区平均透明度110.0 cm、水道平均透明度50.0 cm、东湖区平均透明度80.0 cm;春季西湖区平均透明度72.8 cm、水道平均透明度45.7 cm、东湖区平均透明度80.8 cm,两个湖区在各个季节中透明度均无太大差异。
3.2.1.3 pH值
2020-2021年,全湖平均pH为8.55,其中秋季西湖区平均pH值8.22、水道平均pH值8.65、东湖区平均pH值8.28;冬季西湖区平均pH值9.33、水道平均pH值8.98、东湖区平均pH值8.34;春季西湖区平均pH值8.61、水道平均pH值8.33、东湖区平均pH值8.42,两湖在各个季节中pH值均无太大差异,但冬季西湖区偏弱碱性。
3.2.1.4 电导率
2020-2021年,全湖平均电导率为187.7 µs/cm,其中秋季西湖区平均电导率为150.3 µs/cm、水道平均电导率为169.8 µs/cm、东湖区平均电导率为221.6 µs/cm;冬季西湖区平均电导率为136.7 µs/cm、水道平均电导率为119.7 µs/cm、东湖区平均电导率为259.4 µs/cm;春季西湖区平均电导率为185.9 µs/cm、水道平均电导率为208.5 µs/cm、东湖区平均电导率为237.0 µs/cm。整体上东湖区电导率比西湖区高,表明东湖区离子含量高于西湖区。
3.2.1.5 溶解氧
2020-2021年,全湖平均溶解氧为10.10 mg/L,其中秋季西湖区平均溶解氧为9.25 mg/L、水道平均溶解氧为10.51 mg/L、东湖区平均溶解氧为10.15 mg/L;冬季西湖区平均溶解氧为11.92 mg/L、水道平均溶解氧为9.98 mg/L、东湖区平均溶解氧为11.21 mg/L;春季西湖区平均溶解氧为9.95 mg/L、水道平均溶解氧为8.05 mg/L、东湖区平均溶解氧为8.99mg/L,整体上两个湖区溶解氧无太大差异。
3.2.1.6 浊度
2020-2021年,全湖平均浊度为6.7 NTU,其中秋季西湖区平均浊度为7.1 NTU、水道平均浊度为6.2 NTU、东湖区平均浊度为3.4 NTU;冬季西湖区平均浊度为3.4 NTU、水道平均浊度为11.2 NTU、东湖区平均浊度为3.6 NTU;春季西湖区平均浊度为5.3 NTU、水道平均浊度为16.0 NTU、东湖区平均浊度为4.2 NTU。浊度是由于水中含有泥沙、黏土、有机物、无机物、生物和微生物的悬浮体造成的,浑浊也是水污染的一个重要指标,秋季西湖区浊度比东湖区高,说明西湖区污染程度可能比东湖区严重,其余季节两个湖区浊度无太大差异(表3-3、3-4)。
年份 | 点位 | 水深 (m) | 透明度 (cm) | pH | 溶解氧 (mg·L-1) | 水温 (℃) | 电导率 (us·cm-1) | 浊度 (NTU) |
2 0 2 0 年 11 月 | XLH 1 | 3.4 | 60 | 7.43 | 6.77 | 18.9 | 154.8 | 5.9 |
XLH 2 | 2.5 | 45 | 8.04 | 9.10 | 18.7 | 151.8 | 9.5 | |
XLH 3 | 3.6 | 60 | 8.30 | 9.87 | 18.3 | 149.1 | 6.5 | |
XLH 4 | 3.6 | 50 | 8.51 | 9.99 | 18.4 | 149.4 | 7.3 | |
XLH 5 | 3.3 | 60 | 8.48 | 10.05 | 18.1 | 148.0 | 6.7 | |
XLH 6 | 3.1 | 50 | 8.53 | 9.72 | 18.2 | 148.9 | 6.6 | |
XLH 7 | 2.4 | 50 | 8.77 | 10.43 | 18.1 | 148.7 | 7.8 | |
XLH 8 | 2.8 | 50 | 8.73 | 10.96 | 18.5 | 155.4 | 5.5 | |
XLH 9 | 2.5 | 60 | 8.46 | 10.13 | 18.6 | 205.3 | 5.2 | |
XLH 10 | 3.1 | 60 | 8.64 | 11.20 | 18.5 | 199.1 | 3.6 | |
XLH 11 | 3.1 | 50 | 8.12 | 9.36 | 18.2 | 219.7 | 3.5 | |
XLH 12 | 3.3 | 50 | 8.49 | 10.49 | 18.9 | 214.3 | 3.8 | |
XLH 13 | 3.4 | 60 | 8.45 | 10.71 | 18.7 | 222.4 | 3.3 | |
XLH 14 | 3.2 | 60 | 8.04 | 9.05 | 18.8 | 228.2 | 3.4 | |
XLH 15 | 2.7 | 75 | 7.94 | 10.10 | 18.5 | 245.7 | 2.9 | |
2 0 2 1 年 2 月 | XLH 1 | 1.5 | 85 | 8.95 | 11.96 | 17.7 | 176.6 | 3.9 |
XLH 2 | 1.4 | 90 | 9.03 | 12.17 | 17.6 | 158.8 | 3.8 | |
XLH 3 | 1.7 | 150 | 9.79 | 13.09 | 17.6 | 110.5 | 1.1 | |
XLH 4 | 1.7 | 130 | 9.32 | 12.28 | 18.0 | 130.0 | 1.8 | |
XLH 5 | 1.5 | 90 | 9.33 | 11.11 | 17.5 | 104.2 | 5.8 | |
XLH 6 | 1.3 | 90 | 9.53 | 10.93 | 17.6 | 139.8 | 4.2 | |
XLH 7 | 0.8 | 60 | 8.87 | 9.81 | 17.2 | 105.4 | 8.5 | |
XLH 8 | 0.5 | 30 | 9.15 | 9.73 | 21.4 | 118.0 | 16.8 | |
XLH 9 | 0.5 | 50 | 8.91 | 10.41 | 21.4 | 135.6 | 8.4 | |
XLH 10 | 0.8 | 80 | 8.14 | 9.10 | 20.5 | 295.6 | 3.3 | |
XLH 11 | 0.9 | 90 | 8.21 | 9.07 | 20.4 | 298.3 | 3.9 | |
XLH 12 | 1.2 | 120 | 8.03 | 8.69 | 19.4 | 265.0 | 2.2 | |
XLH 13 | 0.8 | 80 | 8.50 | 12.06 | 19.4 | 233.1 | 3.1 | |
XLH 14 | 0.8 | 80 | 8.90 | 14.68 | 21.5 | 189.1 | 2.5 | |
XLH 15 | 0.5 | 50 | 8.24 | 13.66 | 21.9 | 275.0 | 6.5 | |
2 0 2 1 年 4 月 | XLH 1 | 2.4 | 90 | 8.53 | 9.96 | 25.0 | 192.3 | 3.9 |
XLH 2 | 2.6 | 66 | 8.70 | 10.44 | 24.4 | 189.1 | 4.1 | |
XLH 3 | 2.7 | 80 | 8.47 | 9.70 | 22.9 | 183.8 | 4.8 | |
XLH 4 | 2.6 | 73 | 8.75 | 10.41 | 23.7 | 183.8 | 5.7 | |
XLH 5 | 2.3 | 70 | 8.44 | 9.31 | 22.8 | 183.6 | 7.0 | |
XLH 6 | 2.4 | 58 | 8.74 | 9.89 | 23.9 | 182.6 | 6.2 | |
XLH 7 | 1.6 | 61 | 8.30 | 8.70 | 23.0 | 222.1 | 9.1 | |
XLH 8 | 1.9 | 38 | 8.21 | 7.37 | 23.5 | 199.3 | 19.5 | |
XLH 9 | 1.8 | 38 | 8.47 | 8.08 | 23.3 | 204.0 | 19.5 | |
XLH 10 | 2.2 | 71 | 8.47 | 8.86 | 23.3 | 199.4 | 6.8 | |
XLH 11 | 2.1 | 74 | 8.40 | 9.30 | 23.1 | 217.0 | 4.4 | |
XLH 12 | 2.3 | 90 | 8.47 | 9.13 | 23.6 | 248.6 | 3.7 | |
XLH 13 | 1.8 | 80 | 8.37 | 8.42 | 23.3 | 232.9 | 4.5 | |
XLH 14 | 2.2 | 95 | 8.37 | 8.44 | 24.1 | 258.0 | 3.1 | |
XLH 15 | 2.2 | 75 | 8.44 | 9.81 | 24.6 | 265.8 | 2.9 |
表3-4 2020-2021年西凉湖各水域水体理化特征季节分布
季节 | 湖区 | 水深 (m) | 透明度 (cm) | pH | 溶解氧 (mg L-1) | 水温 (℃) | 电导率 (us cm-1) | 浊度 (NTU) | |
秋 | 西湖区 | 3.3 | 54.2 | 8.22 | 9.25 | 18.4 | 150.3 | 7.1 | |
水道 | 2.6 | 53.3 | 8.65 | 10.51 | 18.4 | 169.8 | 6.2 | ||
东湖区 | 3.1 | 59.2 | 8.28 | 10.15 | 18.6 | 221.6 | 3.4 | ||
冬 | 西湖区 | 1.5 | 110.0 | 9.33 | 11.92 | 17.7 | 136.7 | 3.4 | |
水道 | 0.6 | 50.0 | 8.98 | 9.98 | 20.0 | 119.7 | 11.2 | ||
东湖区 | 0.8 | 80.0 | 8.34 | 11.21 | 20.5 | 259.4 | 3.6 | ||
春 | 西湖区 | 2.5 | 72.8 | 8.61 | 9.95 | 23.8 | 185.9 | 5.3 | |
水道 | 1.8 | 45.7 | 8.33 | 8.05 | 23.3 | 208.5 | 16.0 | ||
东湖区 | 2.1 | 80.8 | 8.42 | 8.99 | 23.7 | 237.0 | 4.2 | ||
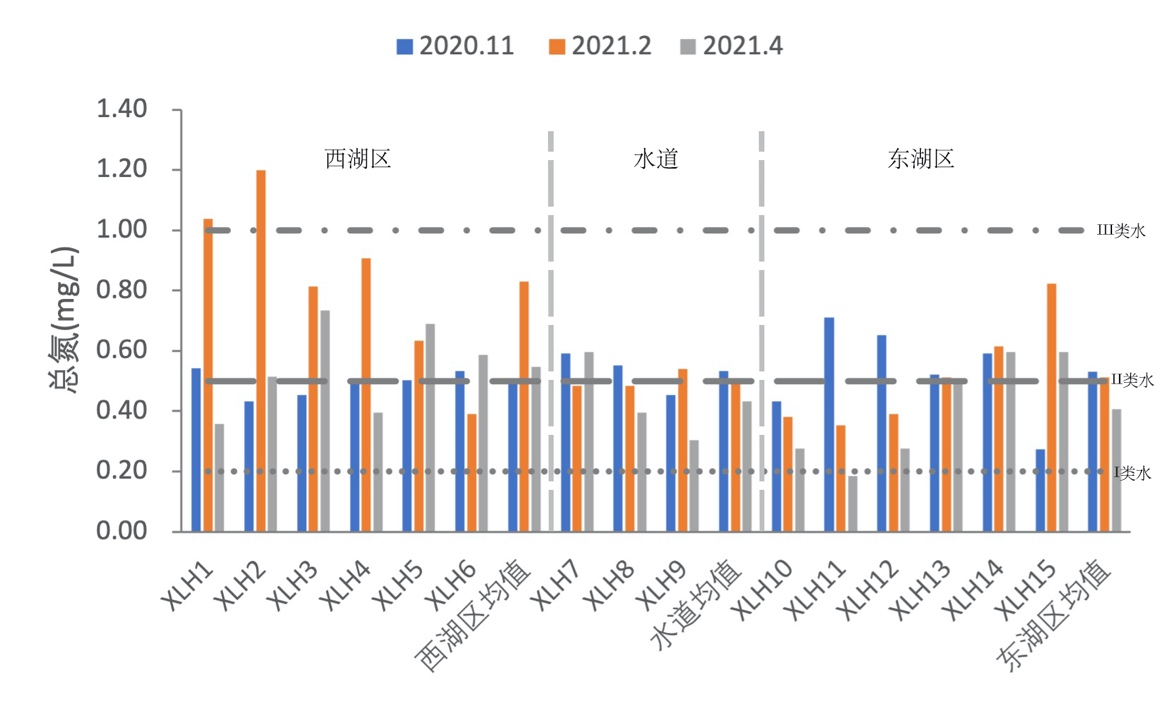
3.2.2.1 总氮
2020-2021年,全湖总氮含量变化范围为0.36-1.20 mg/L,均值为0.62 mg/L;东湖区总氮含量变化范围为0.18-0.82 mg/L,均值为0.48 mg/L;水道总氮含量变化范围为0.30 mg/L-0.60 mg/L,均值为0.49 mg/L。其中秋季均值为0.52 mg/L,冬季均值为0.64 mg/L,春季均值为0.48 mg/L。根据地表水环境质量标准(GB3838-2002),水体总氮基本处于III类水水平,其中西湖区冬季1号、2号区域达到了IV类水水平,其余区域秋、冬、春季处于III类水水平;东湖区处于III类水水平(图3-2)。
秋季西湖区总氮平均值为0.49 mg/L,东湖区总氮平均值为0.53 mg/L,水道总氮平均值为0.54 mg/L,呈现东湖区>水道>西湖区;冬季西湖区总氮平均值为0.83 mg/L,东湖区总氮平均值为0.51 mg/L,水道总氮平均值为0.50 mg/L,呈现西湖区>东湖区>水道;春季西湖区总氮平均值为0.55 mg/L,东湖区总氮平均值为0.41 mg/L,水道总氮平均值为0.43 mg/L,呈现西湖区>水道>东湖区。
综上所述,水体总氮含量处于III类水水平,西湖区冬、春季总氮平均含量均高于东湖区,且在冬季更加明显。
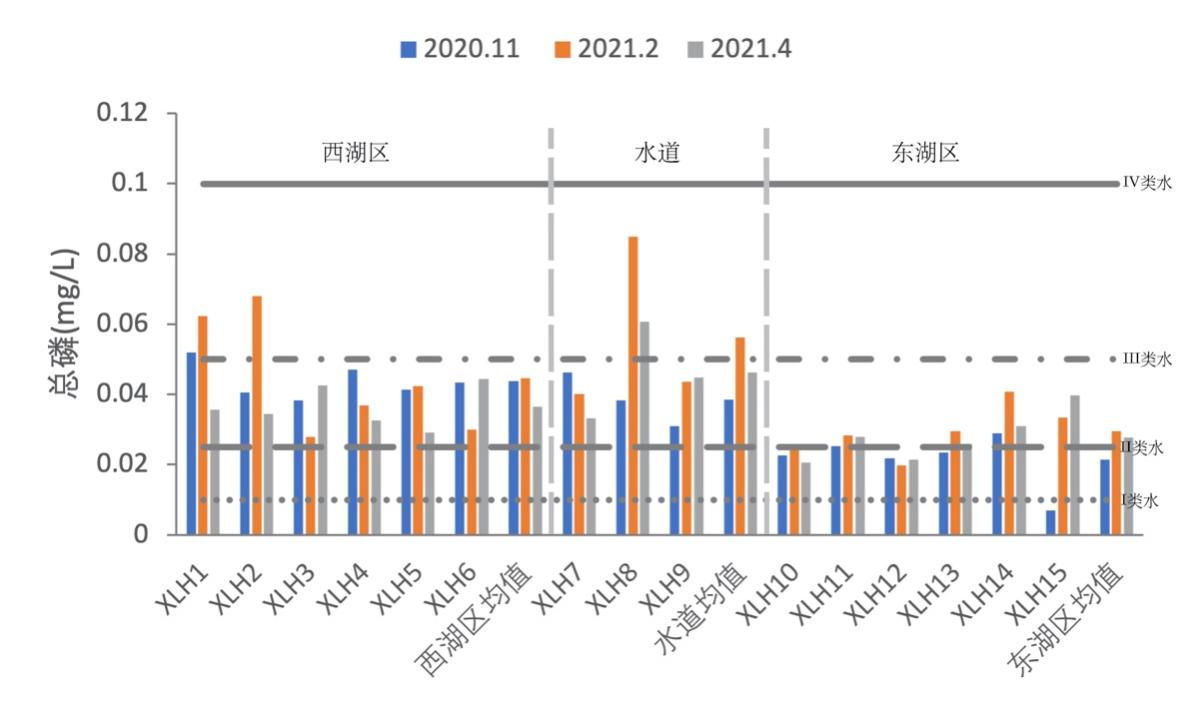
3.2.2.2 总磷
2020-2021年,全湖总磷的含量变化范围为0.0279-0.0681 mg/L,均值为0.0416 mg/L;东湖区总磷含量变化范围为0.0068-0.0408 mg/L,均值为0.0262 mg/L;水道总磷含量变化范围为0.0310-0.0848 mg/L,均值为0.0469 mg/L。其中秋季均值为0.0338 mg/L,冬季均值为0.0408 mg/L,春季均值为0.0349 mg/L。根据地表水环境质量标准(GB3838-2002),2020-2021年水体总磷平均值处于IV类水水平,东湖区处于III类水水平,其中东湖区水在秋季时除XLH14外其余均稳定在II类水水平,但在冬、春季降为III类水水平,表明东湖区水质状况有所变差;西湖区和水道基本处于III类水水平,且西湖区春季水质状况较秋、冬有所好转(图3-3)。
秋季西湖区总磷平均值为0.0438 mg/L,东湖区总磷平均水平为0.0214 mg/L,水道总磷平均水平为0.0385 mg/L,呈现西湖区>水道>东湖区;冬季西湖区总磷平均值为0.0446 mg/L,东湖区总磷平均水平为0.0294 mg/L,水道总磷平均水平为0.0561 mg/L,呈现水道>西湖区>东湖区;春季西湖区总磷平均值为0.0444 mg/L,东湖区总磷平均水平为0.0277 mg/L,水道总磷平均水平为0.0462 mg/L,呈现水道>西湖区>东湖区。
综上所述,西湖区三个季节总磷平均含量均高于东湖区,且在2020-2021年间,西湖区、水道和东湖区总磷变化趋势基本相同。
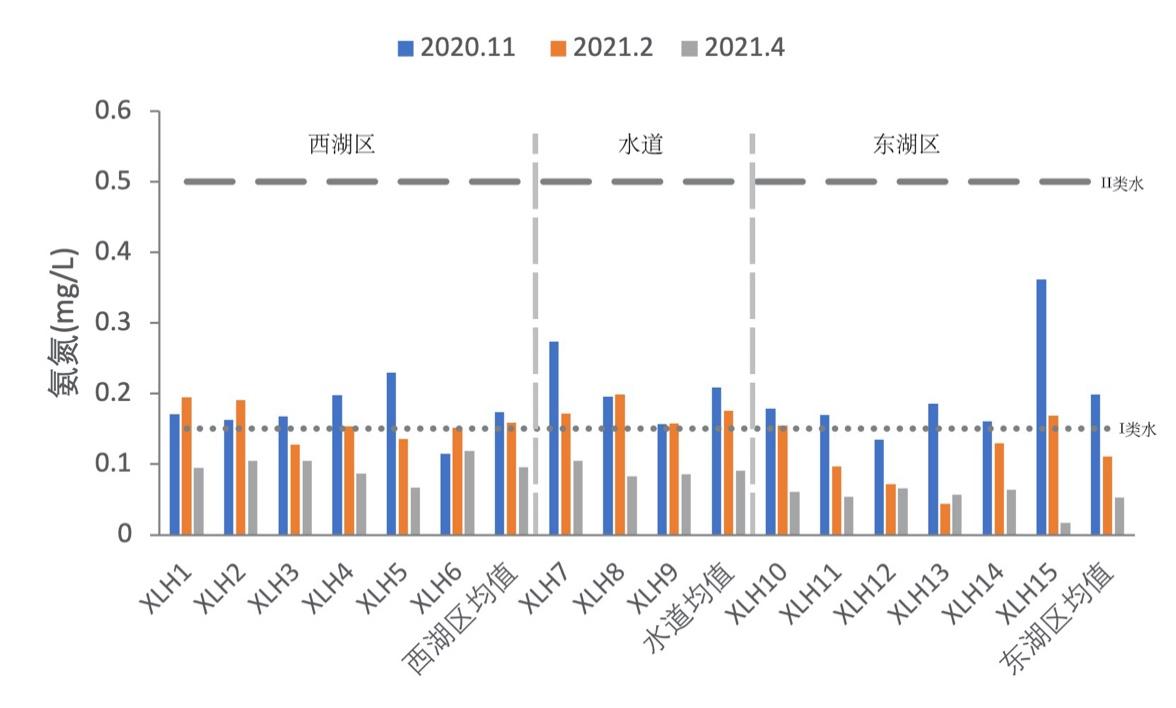
3.2.2.3 氨氮
2020-2021年,西湖区氨氮的含量变化范围为0.0671-0.2299 mg/L,均值为0.1429 mg/L;东湖区氨氮的含量变化范围为0.0167-0.3621 mg/L,均值为0.1288 mg/L;水道氨氮的含量变化范围为0.0829-0.2741 mg/L,均值为0.1588 mg/L。其中秋季均值为0.1901 mg/L,冬季均值为0.1432 mg/L,春季均值为0.0779 mg/L。根据地表水环境质量标准(GB3838-2002),水体氨氮整体处于II类水水平,冬季氨氮含量均达到I类水水平,秋季氨氮含量基本处于II类水平,且东湖区秋季15号区域的含量达到最高(如图3-4)。
秋季西湖区氨氮均值为0.1738 mg/L,东湖区氨氮均值为0.1786 mg/L,水道氨氮均值为0.2089 mg/L,呈现水道>东湖区>西湖区;冬季西湖区氨氮均值为0.1589 mg/L,东湖区氨氮均值为0.1762 mg/L,水道氨氮均值为0.1111 mg/L,呈现水道>东湖区>西湖区;春季西湖区氨氮均值为0.0961 mg/L,东湖区氨氮均值为0.0529 mg/L,水道氨氮均值为0.0913 mg/L,呈现西湖区>水道>东湖区。
综上所述,水体氨氮整体处于II类水水平,相对含量较低。但东、西湖区氨氮含量在三个季节中呈现相同的下降趋势,表明水体氨氮循环逐渐完整,氨氮含量逐渐减少。
3.2.2.4 高锰酸盐指数
2020-2021年,西湖区高锰酸盐的含量变化范围为4.11-6.90 mg/L,均值为5.38 mg/L;东湖区高锰酸盐的含量变化范围为1.69-6.43 mg/L,均值为4.15 mg/L;水道高锰酸盐的含量变化范围为3.93 mg/L-7.29 mg/L,均值为5.19 mg/L。其中秋季均值为5.63 mg/L,冬季均值为4.66 mg/L,春季均值为4.31 mg/L。根据地表水环境质量标准(GB3838-2002),西湖区春季除XLH2、XLH4外其他均处于III类水水平,秋季部分区域为IV类水;东湖区除秋季10号区域外,其余均处于III类水水平;且从西湖区向东湖区水质逐渐从IV类水转为III类水(如图3-5)。
秋季水体高锰酸盐指数均值为6.22 mg/L,东湖区高锰酸盐指数均值为4.77 mg/L,水道高锰酸盐指数均值为6.19 mg/L,呈现西湖区>水道>东湖区;冬季西湖区高锰酸盐指数均值为5.53 mg/L,东湖区高锰酸盐指数均值为3.75 mg/L,水道高锰酸盐指数均值为4.72 mg/L,呈现西湖区>水道>东湖区;春季西湖区高锰酸盐指数均值为4.40 mg/L,东湖区高锰酸盐指数均值为4.04 mg/L,水道高锰酸盐指数均值为4.67 mg/L,呈现水道>西湖区>东湖区。

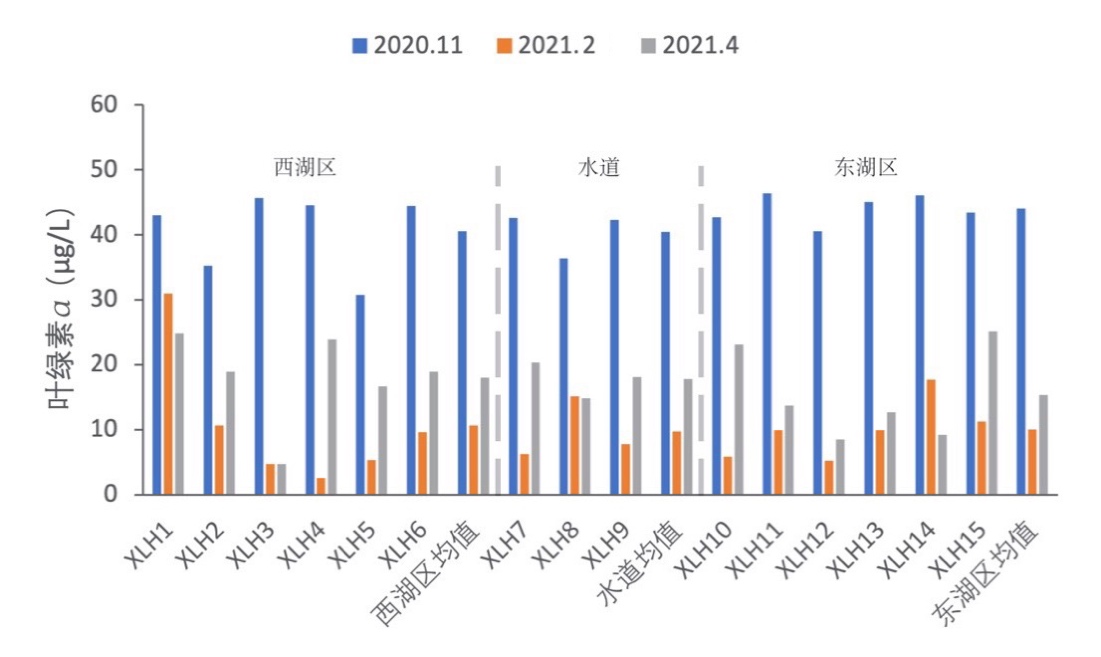
3.2.2.5 叶绿素a
2020-2021年,西湖区叶绿素a变化范围为2.64-45.64 μg/L,均值为23.11 μg/L;东湖区叶绿素a变化范围为5.22-46.42 μg/L,均值为23.16 μg/L;水道叶绿素a变化范围为6.32-42.61 μg/L,均值为22.66 μg/L。总体来看,叶绿素a含量秋季>春季>冬季,东西湖区各季节叶绿素a含量大致相同(图3-6)。
3.2.2.6 富营养化指数
营养程度评价方法采用《地表水环境质量评价技术规范》中推荐的综合营养状态指数(即相关加权营养状态指数TLI)方法,该方法的计算分析方法如下。
综合营养状态指数计算公式:
该方法以修正的卡尔森营养状态指数为基础,以叶绿素a的状态指数为基准,从其余参数的状态指数中,再选择2-3个与基准状态指数比较接近(指绝对值偏差较小)的状态指数,同基准状态指数一起进行相关加权综合。
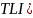
式中:TLI(∑)—综合营养状态指数;
Wj—第j种参数的营养状态指数的相关权重。
TLI(j)—代表第j种参数的营养状态指数。
以Chl-a作为基准参数,则第j种参数的归一化的相关权重计算公式为:
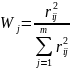
式中:rij—第j种参数与基准参数Chl-a的相关系数;
m—评价参数的个数。
中国湖泊(水库)的Chl-a与其他参数之间的相关关系rij及rij2见表3-5。
表3-5 中国湖泊(水库)部分参数与Chl-a的相关关系rij及rij2值
参数 | Chl-a | TP | TN | SD | CODMn |
rij | 1 | 0.84 | 0.82 | -0.83 | 0.83 |
rij2 | 1 | 0.7056 | 0.6724 | 0.6889 | 0.6889 |
注:参考中国环境监测总站《湖泊(水库)富营养化评价方法及分级技术规定》
单个项目营养状态指数计算公式
TLI(Chl-a)=10(2.5+1.086lnChl-a)
TLI(TP)=10(9.436+1.624lnTP)
TLI(TN)=10(5.453+1.694lnTN)
TLI(SD)=10(5.118-1.94lnSD)
TLI(CODMn)=10(0.109+2.661lnCODMn)
式中:Chl-a单位为mg/m3,SD单位为m;其它项目单位均为mg/L。
湖库营养状态分级
采用0-100的一系列连续数字对湖库营养状态进行分级,包括:贫营养、中营养、富营养、轻度富营养、中度富营养和重度富营养,与污染程度关系如表3-6
营养状态分级 | 评分值TLI(∑) | 定性评价 |
贫营养 | 0<TLI(∑)≤30 | 优 |
中营养 | 30<TLI(∑)≤50 | 良好 |
(轻度)富营养 | 50<TLI(∑)≤60 | 轻度污染 |
(中度)富营养 | 60<TLI(∑)≤70 | 中度污染 |
(重度)富营养 | 70<TLI(∑)≤100 | 重度污染 |
注:参考中国环境监测总站《湖泊(水库)富营养化评价方法及分级技术规定》
运用上述综合营养状态指数法对西凉湖不同区域的水质营养状态进行了计算(表3-7),各个季节综合营养状态指数变动于44.72-53.87之间,均值49.52。水体整体属于轻度富营养化,水质属于轻度污染,水质状况比较差,东湖区基本在中营养化,西湖区综合营养指数均高于东湖区,呈现轻度富营养化(图3-7)。
表3-7 2020-2021年西凉湖各季节不同湖区营养状况综合评价
季节 | 湖区 | TLI | 营养状况 |
秋 | 西湖区 | 53.87 | 轻度富营养 |
水道 | 53.74 | 轻度富营养 | |
东湖区 | 50.55 | 轻度富营养 | |
冬 | 西湖区 | 48.68 | 中营养 |
水道 | 49.74 | 中营养 | |
东湖区 | 44.72 | 中营养 | |
春 | 西湖区 | 48.52 | 中营养 |
水道 | 50.45 | 轻度富营养 | |
东湖区 | 45.55 | 中营养 | |
均值 | 49.52 | 中营养 | |
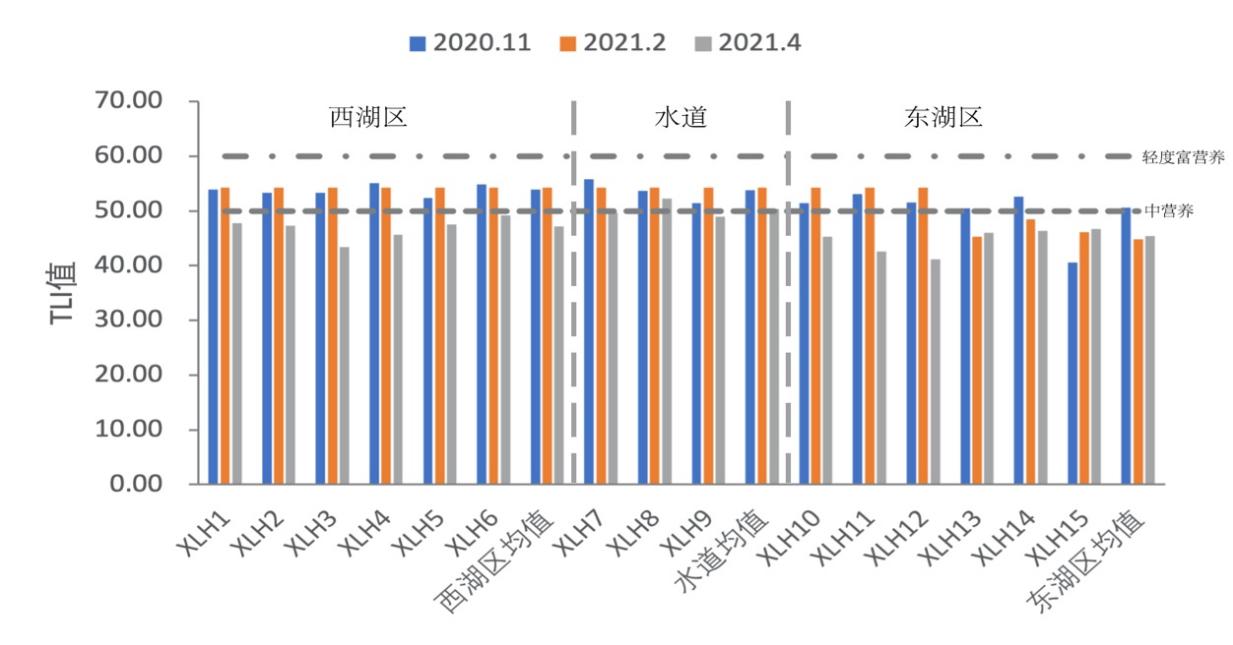
图3-7 2020-2021年西凉湖各季节不同湖区TLI指数
3.2.2.7 水体营养盐综合分析
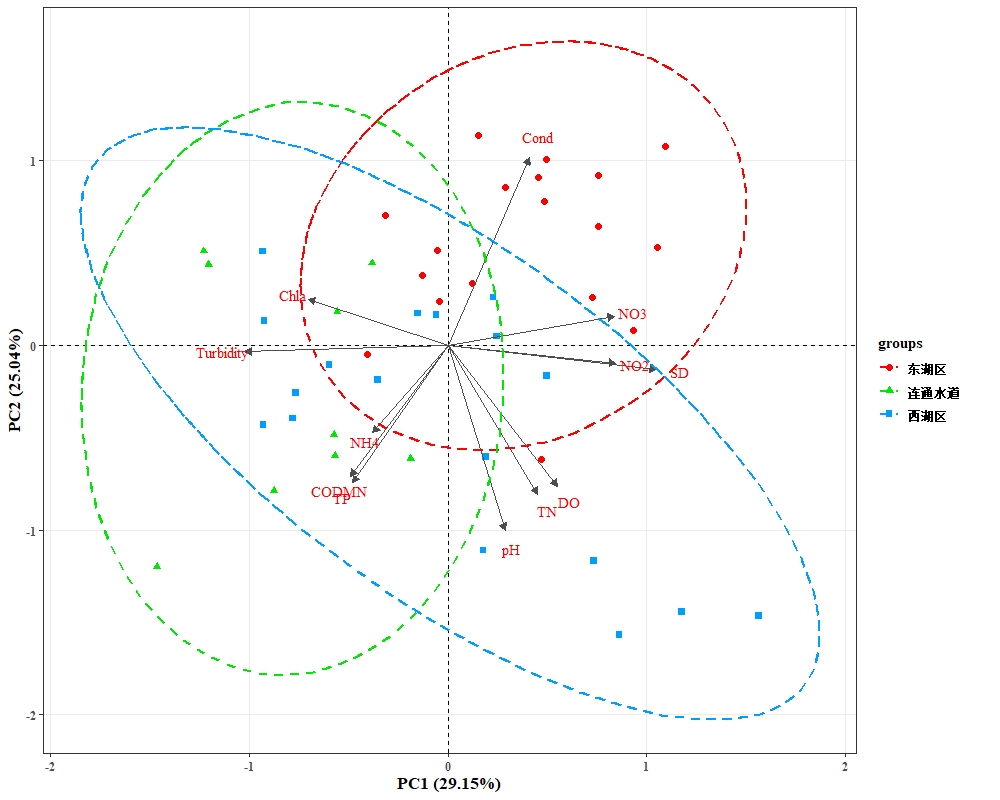
根据水体营养盐指数,对西凉湖2020-2021年水质进行主成分分析(PCA),其中西湖区与连通水道不存在显著性差异(P>0.05),西湖区与东湖区存在显著性差异(P<0.05),东湖区与连通水道存在显著性差异(P<0.05);东湖区主要呈现为电导率、硝酸盐等指标具有相对较高的水平;西湖区和水道主要呈现为叶绿素a、浊度、氨氮、总磷等指标具有相对较高水平(图3-8)。
2017-2020年,对西凉湖水质进行了年际间变化分析,主要水质指标如表3-8。
年份 | 点位 | 总氮 (mg·L-1) | 总磷 (mg·L-1) | 氨氮 (mg·L-1) | 高锰酸盐指数 (mg·L-1) | 叶绿素a (mg·L-1) |
2017 | XLH 1 | 0.83 | 0.0071 | 0.0218 | 4.01 | 4.06 |
XLH 2 | 1.13 | 0.0087 | 0.0295 | 4.18 | 8.44 | |
XLH 3 | 0.75 | 0.0071 | 0.0257 | 4.06 | 6.78 | |
XLH 4 | 1.42 | 0.0055 | 0.0103 | 3.83 | 3.75 | |
XLH 5 | 0.71 | 0.0565 | 0.1061 | 3.42 | 4.84 | |
XLH 6 | 1.12 | 0.0151 | 0.0333 | 4.61 | 4.63 | |
均值 | 0.99 | 0.0166 | 0.0378 | 4.02 | 5.42 | |
2018 | XLH1 | 1.88 | 0.1127 | 0.5657 | 7.53 | 64.78 |
XLH 2 | 1.41 | 0.0671 | 0.4343 | 7.99 | 65.49 | |
XLH 3 | 1.54 | 0.0861 | 1.0366 | 7.38 | 40.88 | |
XLH 4 | 1.56 | 0.0595 | 0.4836 | 6.78 | 35.43 | |
XLH 5 | 2.34 | 0.1355 | 0.5109 | 10.84 | 74.84 | |
XLH 6 | 0.83 | 0.0329 | 0.1495 | 5.51 | 4.13 | |
均值 | 1.59 | 0.0823 | 0.5301 | 7.67 | 47.59 | |
2019 | XLH1 | 1.45 | 0.1388 | 0.0818 | 10.48 | 102.41 |
XLH 2 | 1.22 | 0.1071 | 0.0650 | 10.41 | 81.27 | |
XLH 3 | 1.24 | 0.1190 | 0.0608 | 10.55 | 93.27 | |
XLH 4 | 1.19 | 0.0992 | 0.0608 | 9.44 | 75.23 | |
XLH 5 | 1.29 | 0.1150 | 0.0481 | 10.55 | 97.78 | |
XLH 6 | 1.52 | 0.1388 | 0.0734 | 10.65 | 111.38 | |
均值 | 1.32 | 0.1196 | 0.0650 | 10.35 | 93.56 | |
2020 | XLH1 | 0.54 | 0.0519 | 0.1704 | 5.57 | 43.07 |
XLH 2 | 0.43 | 0.0407 | 0.1627 | 5.96 | 35.27 | |
XLH 3 | 0.45 | 0.0382 | 0.1680 | 6.43 | 45.64 | |
XLH 4 | 0.49 | 0.0471 | 0.1975 | 6.90 | 44.55 | |
XLH 5 | 0.50 | 0.0415 | 0.2300 | 5.96 | 30.75 | |
XLH 6 | 0.53 | 0.0435 | 0.1144 | 6.51 | 44.48 | |
均值 | 0.49 | 0.0438 | 0.1738 | 6.22 | 40.63 |
3.2.3.1 总氮
2017-2020年,全湖总氮变化范围为0.43-2.34 mg/L, 2018年总氮达到最高,但总体上总氮含量在下降,且在2020年达到最低,表明水质在持续改善。总氮含量总体上在地表IV类水, 2018年许多点位达到V类水,其中XLH5达到劣V类水, 2020年处于III类水水平,部分点位处于Ⅱ类水水平。(图3-9)。
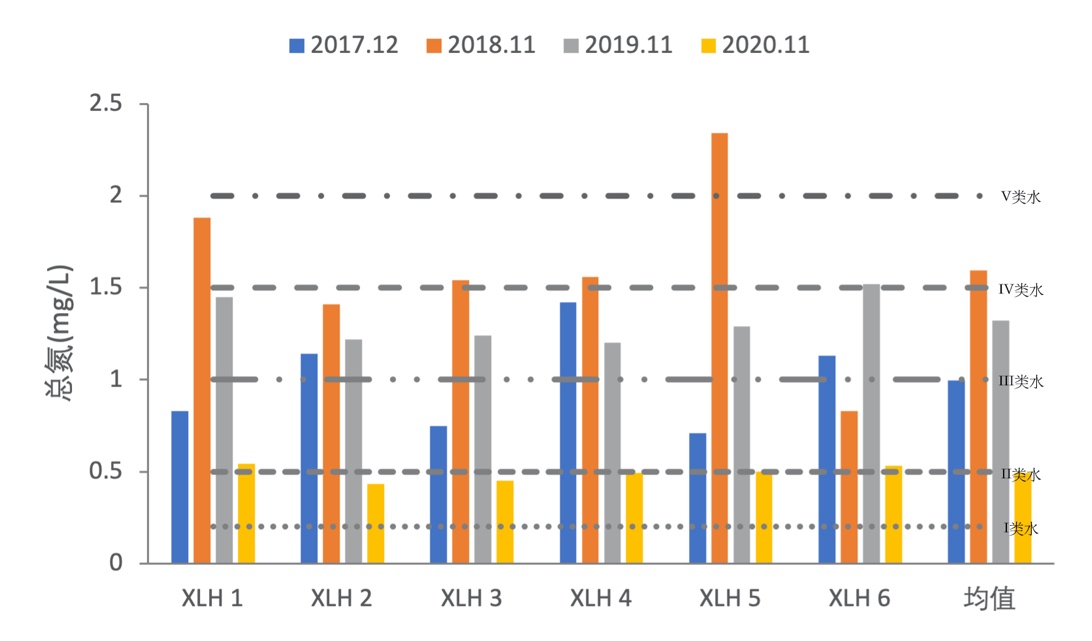
3.2.3.2 总磷
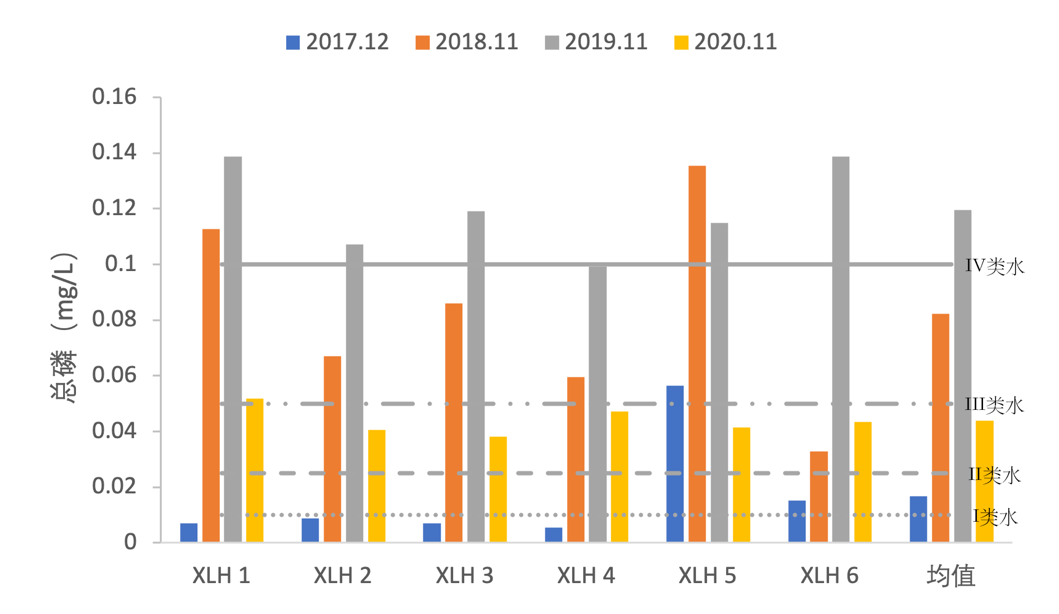
2017-2020年,全湖总磷变化范围为0.0055-0.1388 mg/L, 总磷含量出现变化较大,2017-2019年,总磷含量逐渐增加,但2020年总磷含量有所下降(图3-10),2018年、2019年水质严重恶化,达到了V类水水平,2020年总磷含量有所降低,整体处于III类水水平。
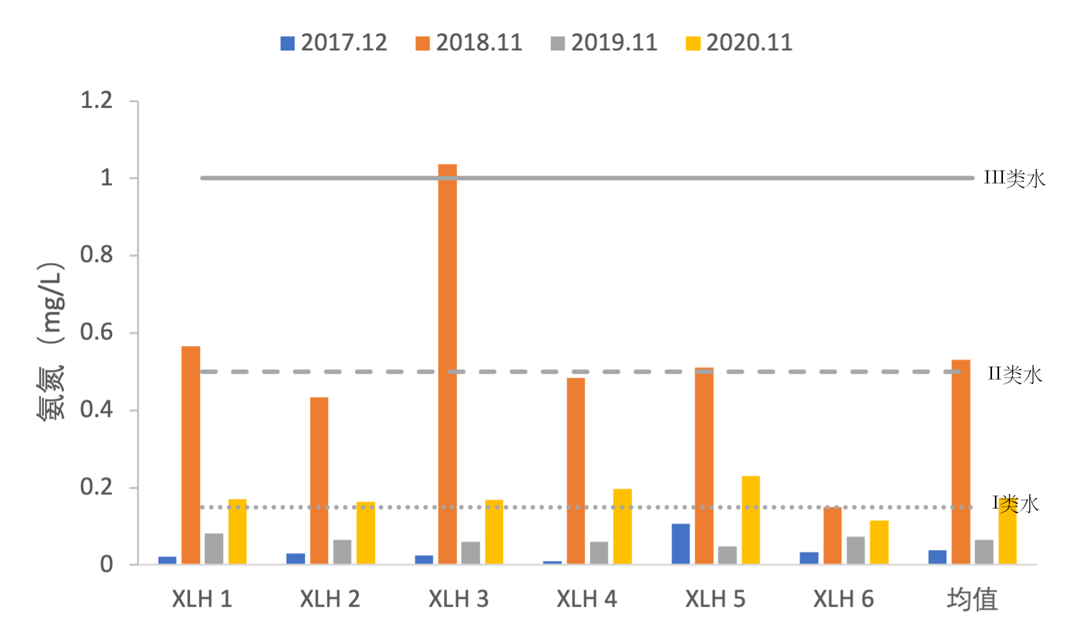
3.2.3.3 氨氮
2017-2020年,全湖氨氮变化范围为0.0103-1.0366 mg/L,氨氮含量除2018年外,总体比较平稳,2018年氨氮达到最大值,为1.0366 mg/L(图3-11),2018年氨氮为III类水水平,XLH3还达到了IV类水水平,到2019年和2020年,氨氮含量降低,逐渐稳定在II类水水平。
3.2.3.4 高锰酸盐指数
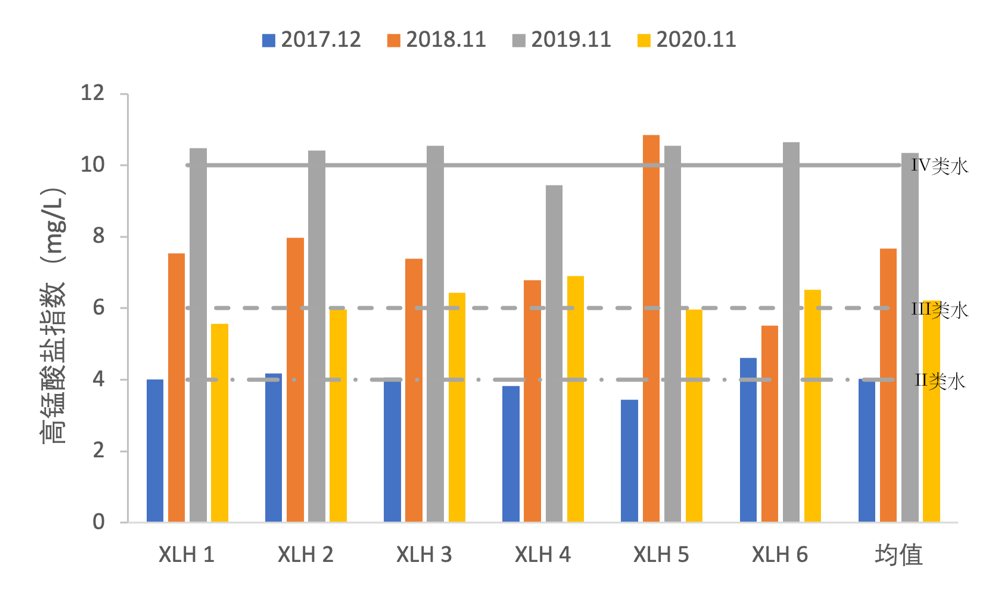
2017-2020年,全湖高锰酸盐指数(CODMn)变化范围为3.44-10.84 mg/L,高锰酸盐指数变动较小,2017-2019年呈上升趋势,并在2019年达到最高,2020年高锰酸盐指数含量有所下降(图3-12)。高锰酸盐指数呈现先上升后下降的趋势,2017年整体处于II类水水平,2019年,高锰酸盐指数达到了V类水水平。
3.2.2.5 叶绿素a
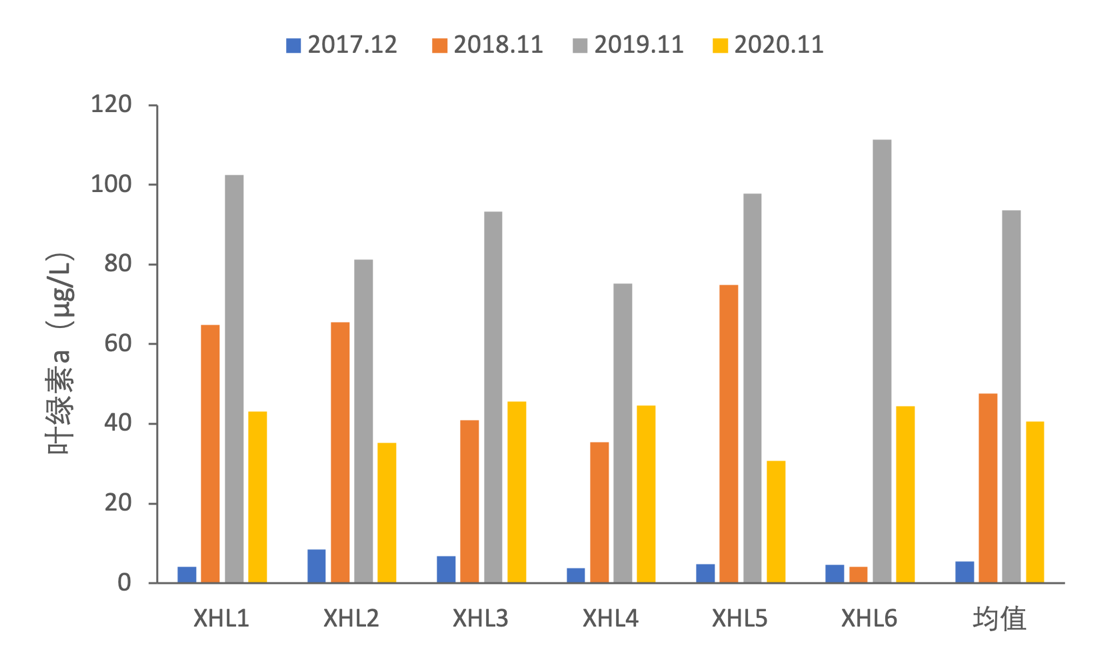
2017-2020年,全湖叶绿素a变化范围为3.74-111.38 μg/L,其中2017年-2019年叶绿素a逐渐增加, 2020年叶绿素a含量有所下降(图3-13)。
3.2.3.6 富营养化指数
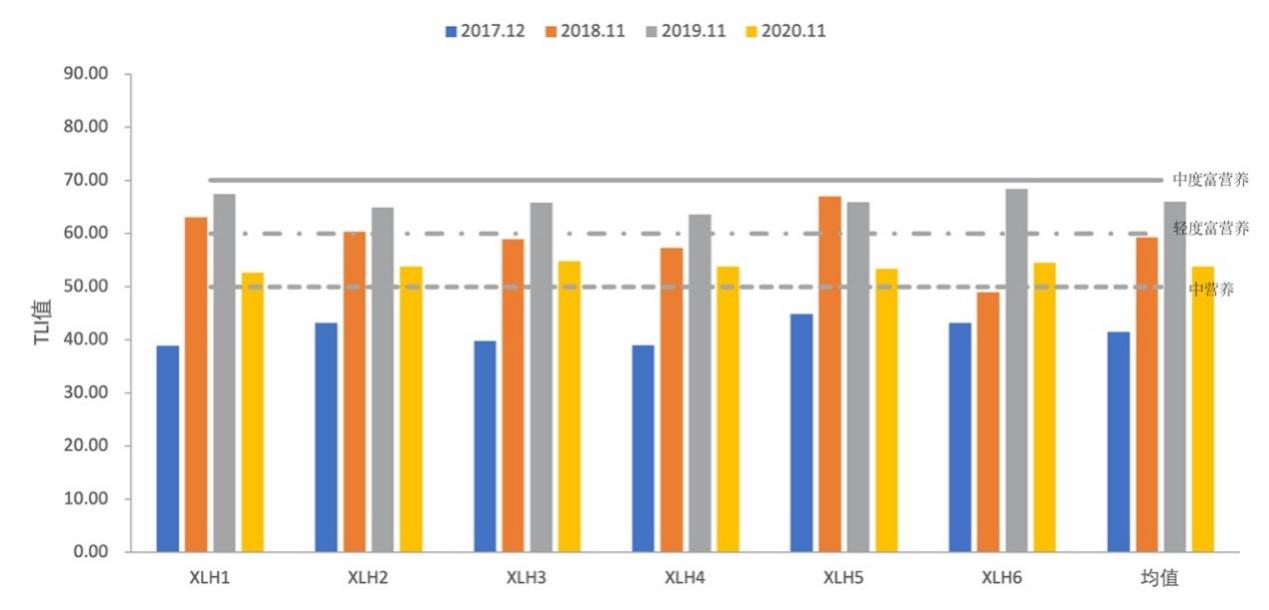
参考3.2.2.5营养指数公式,计算出2017-2020年湖泊富营养化指数(图3-14),其中2017-2019年,湖泊类型从中营养逐渐转变成中度富营养,2020年又变成中营养(表3-9)。
年份 | 湖区 | TLI | 营养 状况 |
2017 | XLH1 | 39.78 | 中营养 |
2017 | XLH2 | 39.67 | 中营养 |
2017 | XLH3 | 44.89 | 中营养 |
2017 | XLH4 | 43.19 | 中营养 |
2017 | XLH5 | 63.04 | 中度富营养 |
2017 | XLH6 | 60.30 | 中度富营养 |
均值 | 48.47 | 中营养 | |
2018 | XLH1 | 58.93 | 轻度富营养 |
2018 | XLH2 | 57.30 | 轻度富营养 |
2018 | XLH3 | 66.93 | 中度富营养 |
2018 | XLH4 | 48.94 | 中营养 |
2018 | XLH5 | 67.43 | 中度富营养 |
2018 | XLH6 | 64.92 | 中度富营养 |
均值 | 60.74 | 中度富营养 | |
2019 | XLH1 | 65.77 | 中度富营养 |
2019 | XLH2 | 63.52 | 中度富营养 |
2019 | XLH3 | 65.92 | 中度富营养 |
2019 | XLH4 | 68.43 | 中度富营养 |
2019 | XLH5 | 52.68 | 轻度富营养 |
2019 | XLH6 | 53.80 | 轻度富营养 |
均值 | 61.68 | 中度富营养 | |
2020 | XLH1 | 54.79 | 轻度富营养 |
2020 | XLH2 | 54.47 | 轻度富营养 |
2020 | XLH3 | 53.33 | 轻度富营养 |
2020 | XLH4 | 55.14 | 轻度富营养 |
2020 | XLH5 | 39.78 | 中营养 |
2020 | XLH6 | 39.67 | 中营养 |
均值 | 49.53 | 中营养 | |
3.2.3.7 水质年际变化综合分析
根据西凉湖水质数据年际变化特点,对2017-2020年水质进行主成分分析(PCA)(图3-15)。其中2017、2018、2019年和2020年每两年之间均存在显著性差异(P<0.05),其中2017年主要表现为透明度较高,2018年和2019年主要表现为氨氮、总氮、总磷、亚硝酸盐、叶绿素a、高锰酸盐指数、浊度等指标较高,2020年则表现为各类营养盐指标均较低。
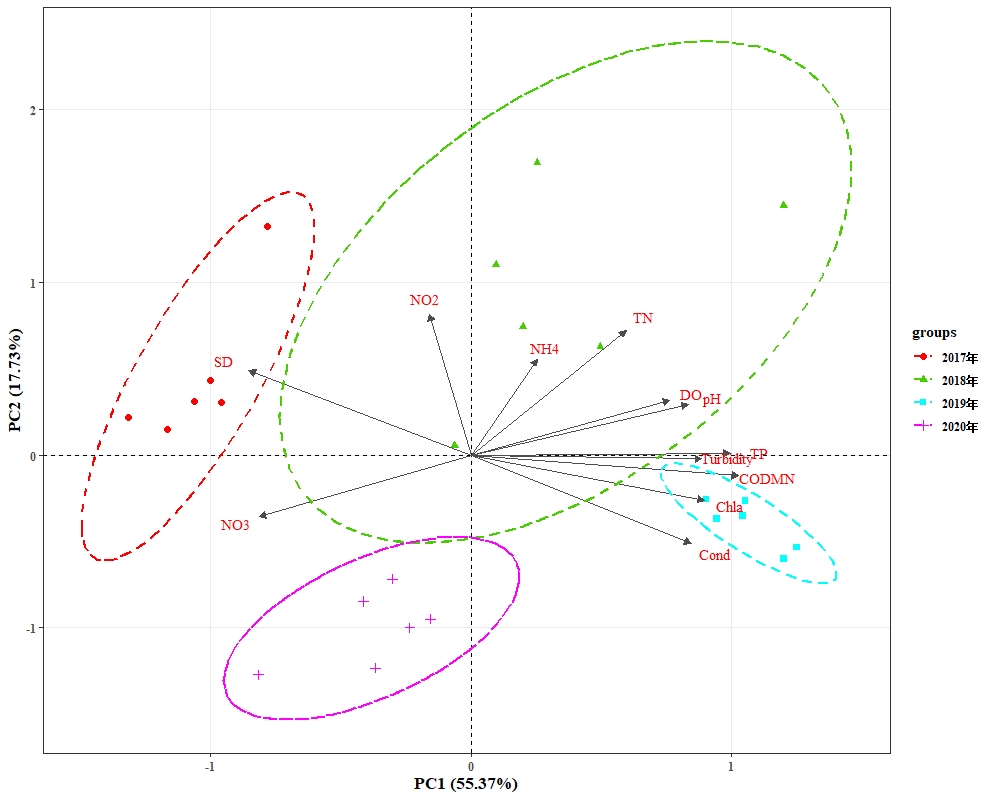
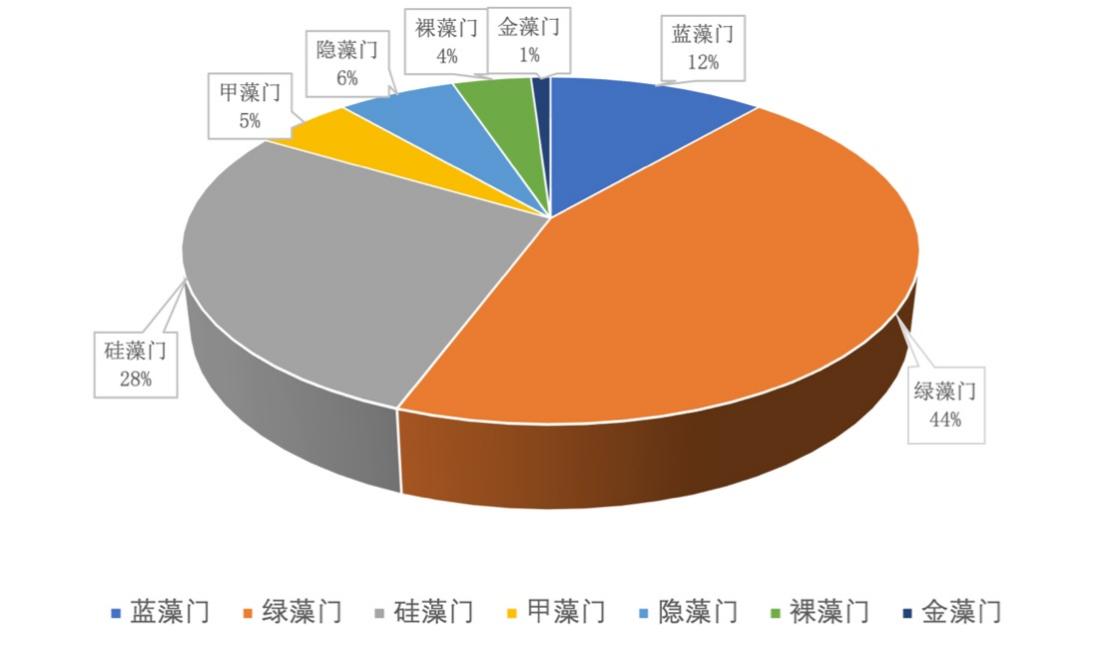
2017-2019年,共镜检出浮游植物7门97种,其中蓝藻门11种、绿藻门43种、硅藻门27种、甲藻门5种、隐藻门6种、裸藻门4种、金藻门1种(表3-10)。由图3-16可得,绿藻门比例最大,占45%,其次是硅藻门占28%、蓝藻门占12%、隐藻门占6%、甲藻门占5%,裸藻门占为4%,金藻门占比例最小为1%。
种类 | 拉丁名 |
蓝藻门 | Cyanophyta |
束丝藻 | Aphanizomenon sp. |
伪鱼腥藻 | Pseudoanabaena sp. |
环圈拟鱼腥藻 | Anabaenopsis circularis |
螺旋鞘丝藻 | Lyngbya contorta |
鱼腥藻 | Anabeana sp. |
微囊藻 | Microcystis sp. |
银灰平裂藻 | Merismopedia glauca |
细小平裂藻 | Merismopedia minima |
弯形小尖头藻 | Raphidiopsis curvata |
细小隐球藻 | Aphanocapsa elachista |
绿色颤藻 | Oscillatoria chlorina |
绿藻门 | Chlorophyta |
叉星鼓藻 | Staurodesmus sp. |
单刺四星藻 | Tetrastrum hastiferum |
单角盘星藻 | Pediastrum simples |
单生卵囊藻 | Oocystis solitaria |
短刺四星藻 | Tetrastrum staurogeniaeforme |
二角盘星藻 | Pediastrum duplex |
二形栅藻 | Scenedesmus dimorphus |
纺锤藻 | Elakatothrix sp. |
浮游角星鼓藻 | Staurastrum planctonicum |
光滑栅藻 | Scenedesmus ecornis |
集星藻 | Actinastrum sp. |
角星鼓藻 | Staurastrum sp. |
具尾四角藻 | Tetraedron caudatum |
空球藻 | Eudorina sp. |
空星藻 | Coelastrum sp. |
孔纹四星藻 | Tetrastrum punctatum |
镰形纤维藻 | Ankistrodesmus falcatus |
卵囊藻 | Oocystis sp. |
卵形衣藻 | Chlamydomonas ovalis |
螺旋弓形藻 | Schroederia spiralis |
螺旋纤维藻 | Ankistrodesmus spiralis |
拟新月藻 | Closteriopsis sp. |
球衣藻 | Chlamydomonas globosa |
三角四角藻 | Tetraedron trigonum |
双眼鼓藻 | Cosmarium bioculatum |
双月藻 | Dicloster sp. |
四刺顶棘藻 | Chodatella quadriseta |
四角盘星藻 | Pediastrum tetras |
四尾栅藻 | Scenedesmus quadricauda |
四足十字藻 | Crucigenia tetrapedia |
蹄形藻 | Kirchneriella sp. |
铜钱形十字藻 | Crucigenia fenestrata |
弯曲栅藻 | Scenedesmus arcuatus |
微小四角藻 | Tetraedron minimum |
纤细角星鼓藻 | Staurastrum gracile |
纤细新月藻 | Closterium gracile |
小球藻 | Chlorella sp. |
小形卵囊藻 | Oocystis parva |
硬弓形藻 | Schroederia robust |
圆鼓藻 | Cosmarium circulare |
月牙藻 | Selenastrum bibraianum |
转板藻 | Mougeotia sp. |
锥顶十字藻 | Crucigenia apiculata |
硅藻门 | Bacillariophyta |
扁圆卵形藻 | Cocconeis placentula |
变异直链藻 | Melosira varians |
粗壮双菱藻 | Surirella robusta |
短小曲壳藻 | Achnanthes exigua |
短小舟形藻 | Navicula exigua |
钝脆杆藻 | Fragilaria capucina |
弧形峨眉藻 | Ceratoneis arcus |
尖布纹藻 | Gyrosigma acuminatum |
尖针杆藻 | Synedra acus |
颗粒直链藻 | Melosira granulata |
菱形藻 | Nitzschia sp. |
扭曲小环藻 | Cyclotella comta |
披针形舟形藻 | Navicula lanceolata |
偏肿桥弯藻 | Cymbella naviculiformis |
普通等片藻 | Diatoma vulgare |
双头舟形藻 | Navicula dicephala |
细菱形藻 | Nitzschia acicularis |
细小桥弯藻 | Cymbella gracilis |
纤细桥弯藻 | Cymbella cuspidate |
线形舟形藻 | Navicula graciloides |
小环藻 | Cyclotella sp. |
小头菱形藻 | Nitzschia microcephala |
缢缩异极藻 | Gomphonema constrictum |
优美桥弯藻 | Cymbella delicatula |
羽纹藻 | Pinnularia sp. |
窄异极藻 | Gomphonema angustatum |
肘状针杆藻 | Synedra ulna |
甲藻门 | Pyrrophyta |
多甲藻 | Peridinium sp. |
飞燕角甲藻 | Ceratium hirundinella |
加顿多甲藻 | Peridinium gatunense |
拟多甲藻 | Peridiniopsis sp. |
微小多甲藻 | Peridinium pusillum |
隐藻门 | Cryptophyta |
倒卵隐藻 | Cryptomonas obovata |
反曲弯隐藻 | Campylomonas reflexa |
具尾逗隐藻 | Komma caudata |
蓝隐藻 | Chroomonas sp. |
马索隐藻 | Cryptomonas marssonii |
细小隐藻 | Cryptomonas tenuis |
裸藻门 | Euglenophyta |
扁裸藻 | Phacus sp. |
尖尾裸藻 | Euglena oxyuris |
囊裸藻 | Trachelomonas sp. |
长尾扁裸藻 | Phacus longicauda |
金藻门 锥囊藻 | Chrysophyta Dinobryon sp. |
总计 | 97 |
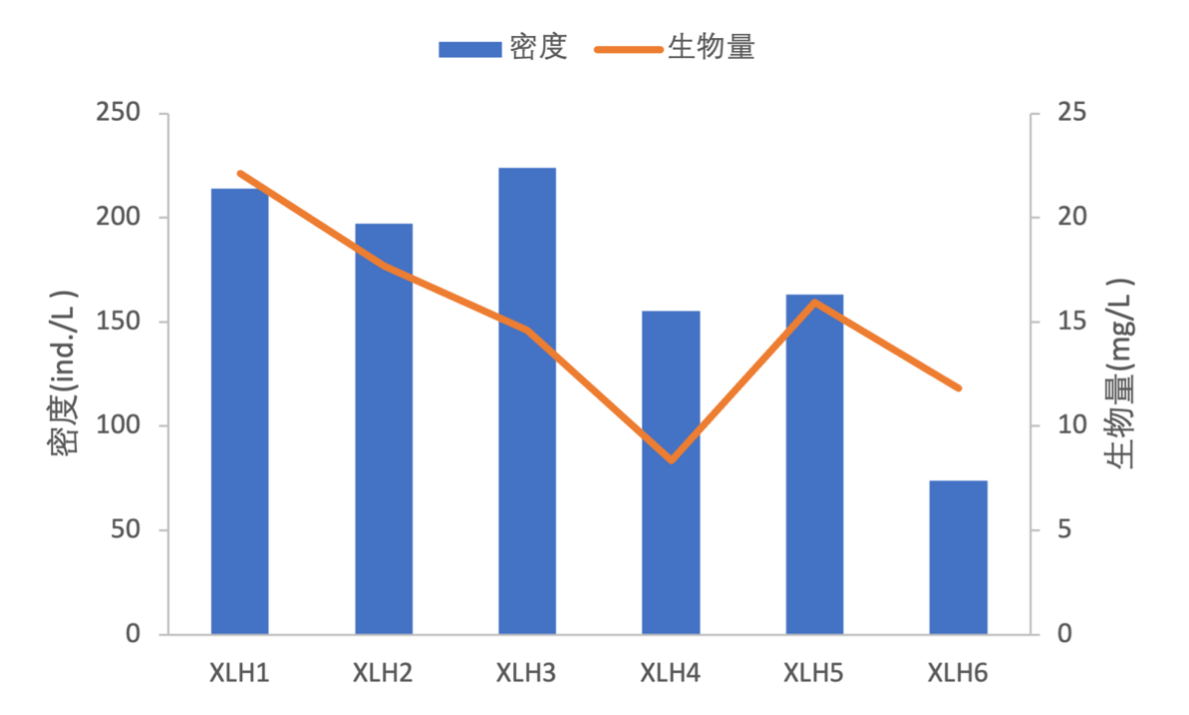
2017-2019年,浮游植物密度变化范围为73.85×106 ind./L-223.96×106 ind./L,平均密度为171.19×106 ind./L,其中XLH3密度最高为223.96×106 ind./L,XLH6最低为73.85×106 ind./L。
蓝藻门密度最高为142.74×106 ind./L,其次为硅藻门为15.54×106 ind./L,金藻门密度最低100.00 ind./L,且仅出现在XLH4(表3-10)。
浮游植物生物量变化范围为8.3387 mg/L-22.1247 mg/L,平均生物量为15.0831 mg/L,其中XLH1生物量最高为22.1247 mg/L,且各藻门生物量也相对较高,其次是XLH2为17.6847 mg/L,XLH4生物量最低为8.3387 mg/L;硅藻门生物量最高为5.5233 mg/L,其次为蓝藻门为3.8356 mg/L,金藻门最低为0.0001 mg/L(表3-11)。
浮游植物密度从XLH1-XLH6呈现有下降的趋势,生物量从XLH1-XLH4呈下降趋势,XLH2、XLH3浮游植物密度较高,藻类爆发的概率也有所增加(图3-17)。
点位 | 蓝藻门 | 绿藻门 | 硅藻门 | 隐藻门 | 甲藻门 | 裸藻门 | 金藻门 | 总计 |
XLH1 | 174.93 | 16.42 | 17.45 | 4.62 | 0.15 | 0.33 | 0.00 | 213.90 |
XLH2 | 165.49 | 8.95 | 20.55 | 1.89 | 0.02 | 0.27 | 0.00 | 197.17 |
XLH3 | 200.47 | 8.50 | 12.52 | 2.32 | 0.02 | 0.13 | 0.00 | 223.96 |
XLH4 | 133.15 | 8.97 | 12.08 | 0.98 | 0.00 | 0.06 | 0.0005 | 155.06 |
XLH5 | 135.03 | 11.39 | 14.64 | 1.86 | 0.08 | 0.04 | 0.00 | 163.04 |
XLH6 | 47.34 | 7.28 | 15.99 | 3.22 | 0.02 | 0.00 | 0.00 | 73.85 |
均值 | 142.74 | 10.25 | 15.54 | 2.48 | 0.05 | 0.14 | 0.0001 | 171.16 |
注:单位(106 cells/L)
点位 | 蓝藻门 | 绿藻门 | 硅藻门 | 隐藻门 | 甲藻门 | 裸藻门 | 金藻门 | 总计 |
XLH1 | 3.9629 | 0.9771 | 5.6406 | 5.3635 | 1.9114 | 4.2693 | 0.0000 | 22.1247 |
XLH2 | 4.8889 | 0.9025 | 6.4737 | 1.0839 | 0.3445 | 3.9913 | 0.0000 | 17.6847 |
XLH3 | 6.1088 | 0.4878 | 4.3615 | 1.1122 | 0.8216 | 1.7101 | 0.0000 | 14.6020 |
XLH4 | 2.6353 | 0.6001 | 3.6941 | 0.0000 | 0.8127 | 0.2437 | 0.0004 | 8.3387 |
XLH5 | 3.9214 | 0.8635 | 5.4880 | 2.9244 | 2.5342 | 0.1963 | 0.0000 | 15.9278 |
XLH6 | 1.4964 | 0.5095 | 7.4819 | 0.0359 | 2.2974 | 0.0000 | 0.0000 | 11.8211 |
均值 | 3.8356 | 0.7234 | 5.5233 | 1.7533 | 1.4536 | 1.7351 | 0.0001 | 15.0832 |
注:单位(mg/L)
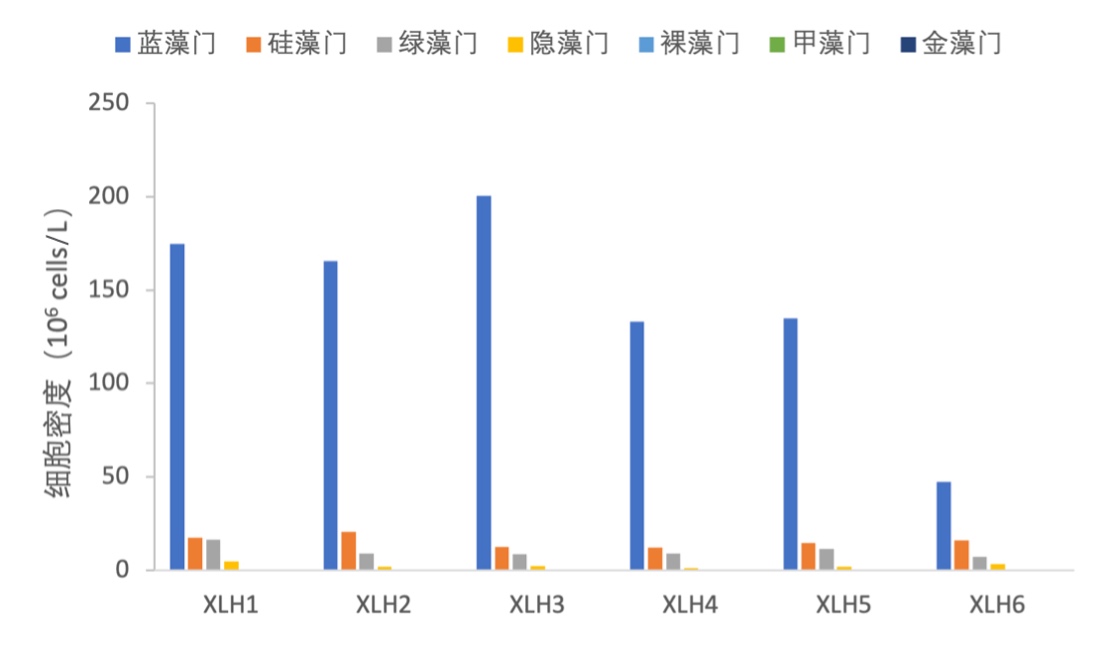
2017-2019年,浮游植物主要以蓝藻门为优势种,硅藻门和绿藻门细胞密度相差不大;在XLH3,蓝藻门具有绝对的优势。硅藻门在XLH6细胞密度有所增加,但优势种主要还是以蓝藻门为主(图3-18和图3-19)。
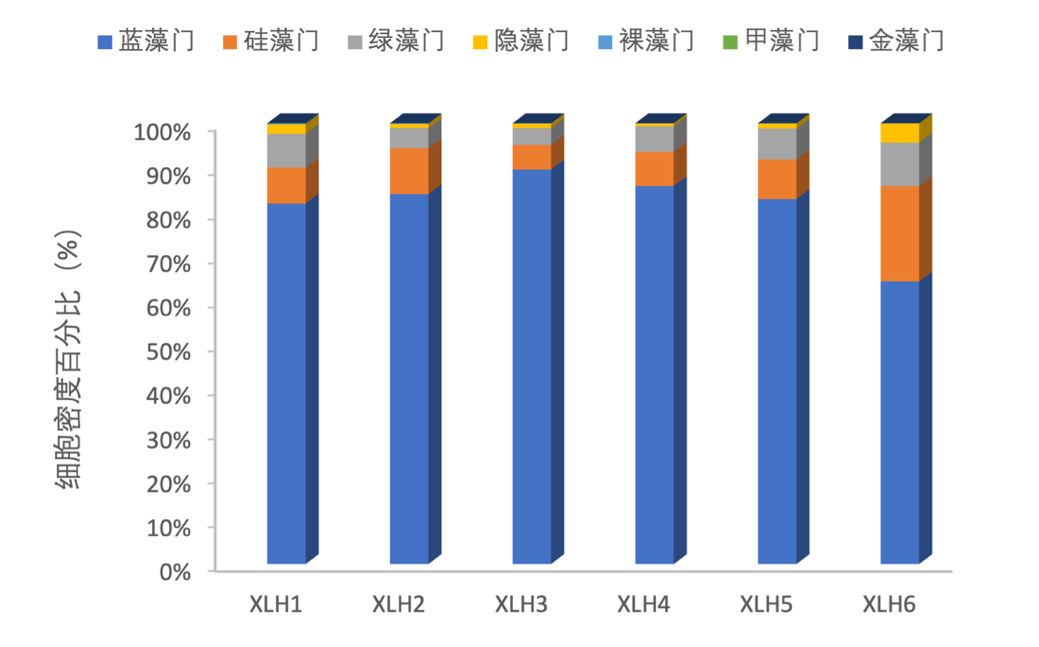
图3-18 2017-2019年西凉湖浮游植物密度组成百分比
2017-2019年,硅藻门占较大的生物量,其次是蓝藻门 ,硅藻门在XLH6生物量最大,XLH1各门类生物量较为平均,不存在明显的优势种,XLH3蓝藻门生物量最大(图3-20、图3-21)。
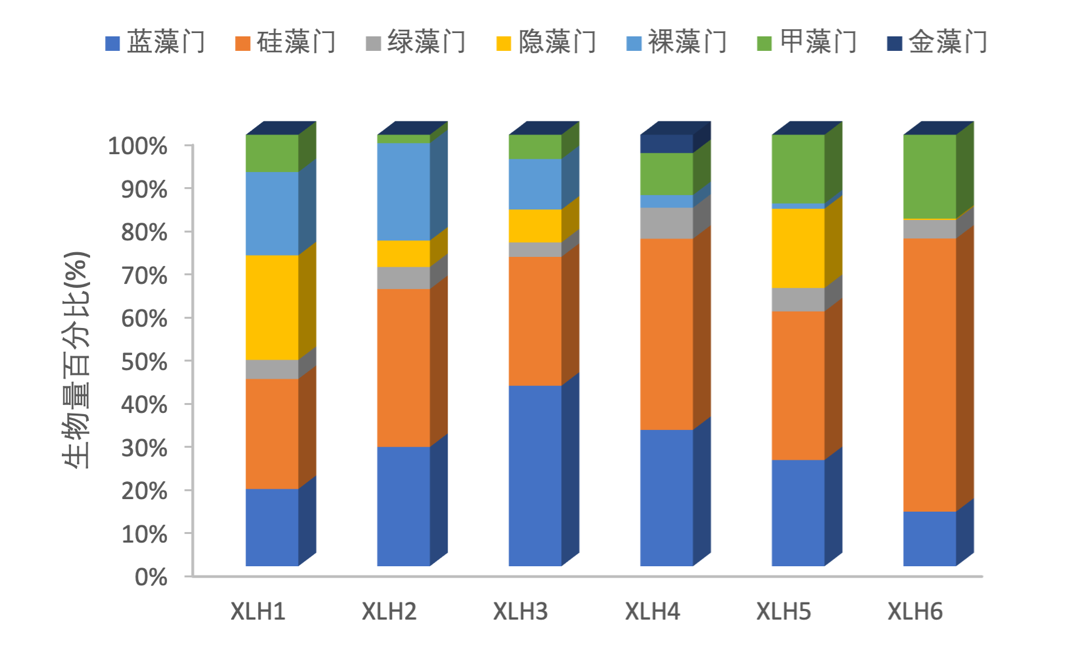
图3-20 2017-2019西凉湖浮游植物生物量组成百分比
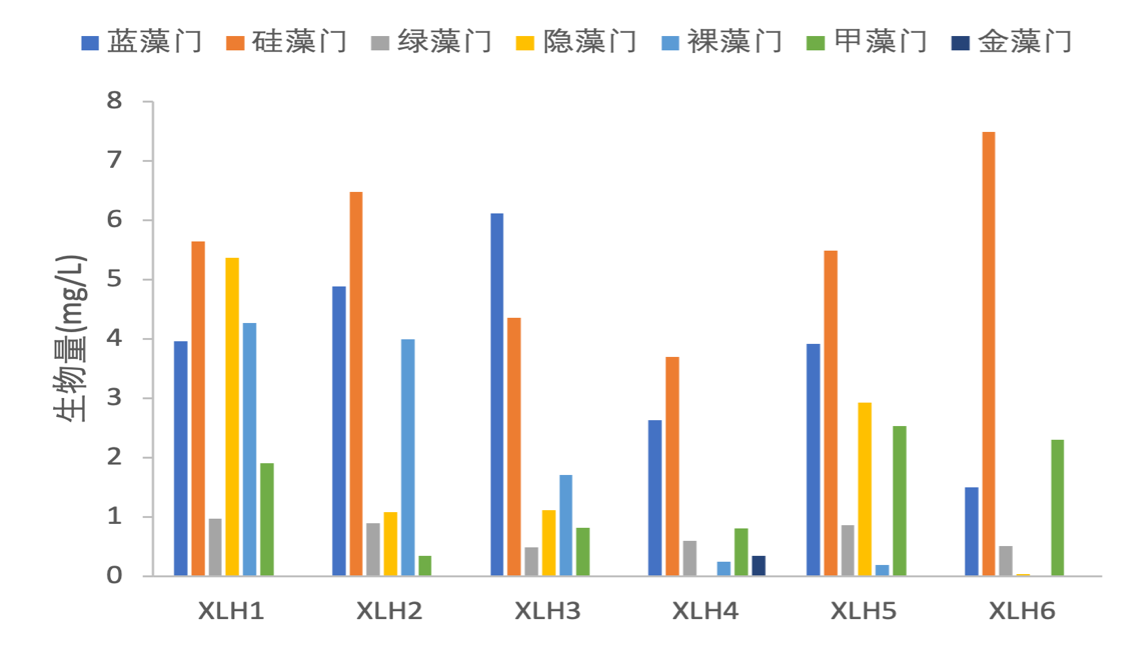
图3-21 2017-2019西凉湖浮游植物生物量组成百分比
2017-2019年,取各个采样点相对密度前十的物种进行排序,且相对密度超过10%的为优势种,西凉湖6个采样点均以蓝藻门的种类优势度最大,束丝藻在XLH1(24.97%)、XLH2(59.08%)、XLH3(64.15%)、XLH4(32.71%)、XLH5(50.82%)为最具优势的物种(表3-13),但在XLH6中相对密度较低。伪鱼腥藻在每个采样点中都为优势种。
种类(41种) | XLH1 | 种类(36种) | XLH2 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
束丝藻 | 24.97% | + | 束丝藻 | 59.08% | + |
伪鱼腥藻 | 22.30% | + | 伪鱼腥藻 | 16.33% | + |
细小平裂藻 | 16.38% | + | 颗粒直链藻 | 7.47% | - |
鱼腥藻 | 10.19% | + | 螺旋鞘丝藻 | 5.76% | - |
微囊藻 | 5.68% | - | 细小平裂藻 | 4.82% | - |
颗粒直链藻 | 5.54% | - | 四尾栅藻 | 1.90% | - |
螺旋鞘丝藻 | 5.22% | - | 微囊藻 | 1.71% | - |
四尾栅藻 | 4.34% | - | 银灰平裂藻 | 1.70% | - |
银灰平裂藻 | 4.07% | - | 马索隐藻 | 0.71% | - |
空星藻 | 1.29% | - | 具尾逗隐藻 | 0.54% | - |
种类(45种) | XLH3 | 种类(35种) | XLH4 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
束丝藻 | 64.15% | + | 束丝藻 | 32.71% | + |
伪鱼腥藻 | 18.44% | + | 伪鱼腥藻 | 30.28% | + |
颗粒直链藻 | 4.22% | - | 银灰平裂藻 | 8.34% | - |
细小平裂藻 | 3.69% | - | 细小平裂藻 | 7.29% | - |
螺旋鞘丝藻 | 2.37% | - | 螺旋鞘丝藻 | 6.61% | - |
银灰平裂藻 | 2.14% | - | 颗粒直链藻 | 5.32% | - |
微囊藻 | 1.46% | - | 微囊藻 | 4.33% | - |
鱼腥藻 | 1.39% | - | 鱼腥藻 | 2.31% | - |
四尾栅藻 | 1.35% | - | 空星藻 | 1.86% | - |
螺旋纤维藻 | 0.77% | - | 四尾栅藻 | 0.94% | - |
种类(38种) | XLH5 | 种类(39种) | XLH6 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
束丝藻 | 50.82% | + | 螺旋鞘丝藻 | 22.09% | + |
伪鱼腥藻 | 13.64% | + | 伪鱼腥藻 | 19.16% | + |
螺旋鞘丝藻 | 8.66% | - | 颗粒直链藻 | 18.38% | + |
颗粒直链藻 | 8.54% | - | 束丝藻 | 12.76% | + |
细小平裂藻 | 7.28% | - | 鱼腥藻 | 9.85% | - |
微囊藻 | 4.65% | - | 微囊藻 | 8.45% | - |
鱼腥藻 | 2.32% | - | 四尾栅藻 | 4.15% | - |
空星藻 | 1.95% | - | 空星藻 | 2.07% | - |
四尾栅藻 | 1.56% | - | 具尾逗隐藻 | 1.97% | - |
单角盘星藻 | 0.59% | - | 倒卵隐藻 | 1.11% | - |
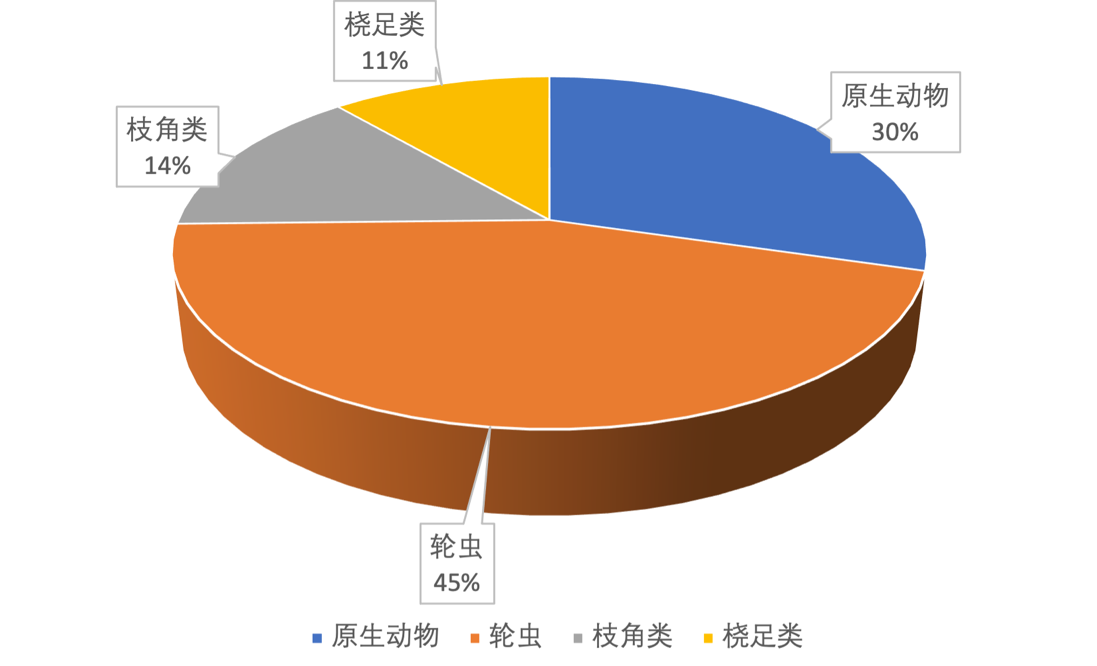
2017-2019年,共镜检出浮游动物71种,原生动物21种,轮虫32种,枝角类10种,桡足类8种(表3-14)。其中,轮虫比例最高为45%,其次是原生动物30%、枝角类14%、桡足类11%(图3-22)。
种类 | 拉丁名 | ||
原生动物 | Protozoa | ||
球砂壳虫 | Difflugia globulosa | ||
岔口砂壳虫 | Difflugia gramen | ||
毛板壳虫 | Coleps hirtus | ||
尾草履虫 | Paramecium caudatum | ||
放射矛刺虫 | Hastatella radians | ||
光明舟形虫 | Lembadion lucens | ||
累枝虫 | Epistylis sp. | ||
急游虫 | Strombidium sp. | ||
裸口虫 | Holophrya sp. | ||
侠盗虫 | Strobilidium sp. | ||
喇叭虫 | Stentor sp. | ||
球形砂壳虫 | Difflugia globulosa | ||
天鹅长吻虫 | Lacrymaria olor | ||
绿急游虫 | Strombidium viride | ||
王氏似铃壳虫 | Tintinnopsis wangi | ||
纤毛虫 | Ciliophora sp. | ||
小单环栉毛虫 | Didinium balbianii nanum | ||
游仆虫 | Euplotes sp. | ||
钟虫 | Vorticella sp. | ||
银白前口虫 | Frontonia leucas | ||
旋回侠盗虫 | Strobilidium gyrans | ||
轮虫 | Rotifera | ||
胶鞘轮虫 | Collotheca sp. | ||
顶生三肢轮虫 | Filinia terminalis | ||
角突臂尾轮虫 | Brachionus angularia | ||
萼花臂尾轮虫 | Brachionus calyciflorus | ||
暗小异尾轮虫 | Trichocerca pusilla | ||
卵形彩胃轮虫 | Chromogaster ovalis | ||
裂痕龟纹轮虫 | Anuraeopsis fissa | ||
裂足轮虫 | Schizocerca diversicornis | ||
锯齿龟纹轮虫 | Anuraeopsis coelata | ||
裂痕龟纹轮虫 | Anuraeopsis fissa | ||
东方角突臂尾轮虫 | Brachionus angularis orientalis | ||
蒲达臂尾轮虫 | Brachionus budapestiensis | ||
短棘剪形臂尾轮虫 | Brachionus forficula reducta | ||
螺形龟甲轮虫 | Keratella cochlearis | ||
广布多肢轮虫 | Polyarthra vulgaris | ||
巨头轮虫 | Cephalodella sp. | ||
罗氏异尾轮虫 | Trichocerca rousseleti | ||
等刺异尾轮虫 | Trichocerca similis | ||
扁平泡轮虫 | Pompholyx complanta | ||
多态胶鞘轮虫 | Collotheca ambigua | ||
小三肢轮虫 | Filinia minuta | ||
无尾无柄轮虫 | Ascomorpha ecaudis | ||
无棘龟甲轮虫 | Keratella tecta | ||
曲腿龟甲轮虫 | Keratella valga | ||
椎尾水轮虫 | Epiphanes senta | ||
前节晶囊轮虫 | Asplanchna priodonta | ||
长肢多肢轮虫 | Polyarthra dolichoptera | ||
梳状疣毛轮虫 | Synchaeta pectinata | ||
疣毛轮虫 | Synchaeta sp. | ||
圆筒异尾轮虫 | Trichocerca cylindrica | ||
鼠异尾轮虫 | Trichocerca rattus | ||
长三肢轮虫 | Filinia longiseta | ||
枝角类 | Cladocera | ||
裸腹溞 | Moina sp. | ||
简弧象鼻溞 | Bosmina coregoni | ||
矩形尖额溞 | Alona rectangula | ||
圆形盘肠溞 | Chydorus sphaericus | ||
里氏薄皮溞 | Leptodora richardi | ||
短尾秀体溞 | Diaphanosoma brachyurum | ||
模糊秀体溞 | Diaphanosoma dubium | ||
脆弱象鼻溞 | Bosmina fatalis | ||
圆形盘肠溞 | Chydorus sphaericus | ||
龟状笔纹溞 | Graptoleberis testudinaria | ||
桡足类 | Copepoda | ||
近邻剑水蚤 | Cyclops vicinus | ||
哲水蚤桡足幼体 | Canaloida Copepodid | ||
如意真剑水蚤 | Eucyclops speratus | ||
剑水蚤桡足幼体 | Cyclopoida Copepodid | ||
跨立小剑水蚤 | Microcyclops (Microcyclops) varicans | ||
刘氏中剑水蚤 | Mesocyclops leuckarti | ||
台湾温剑水蚤 | Thermocyclops taihokuensis | ||
无节幼体 | Nauplius | ||
总计 | 71 | ||
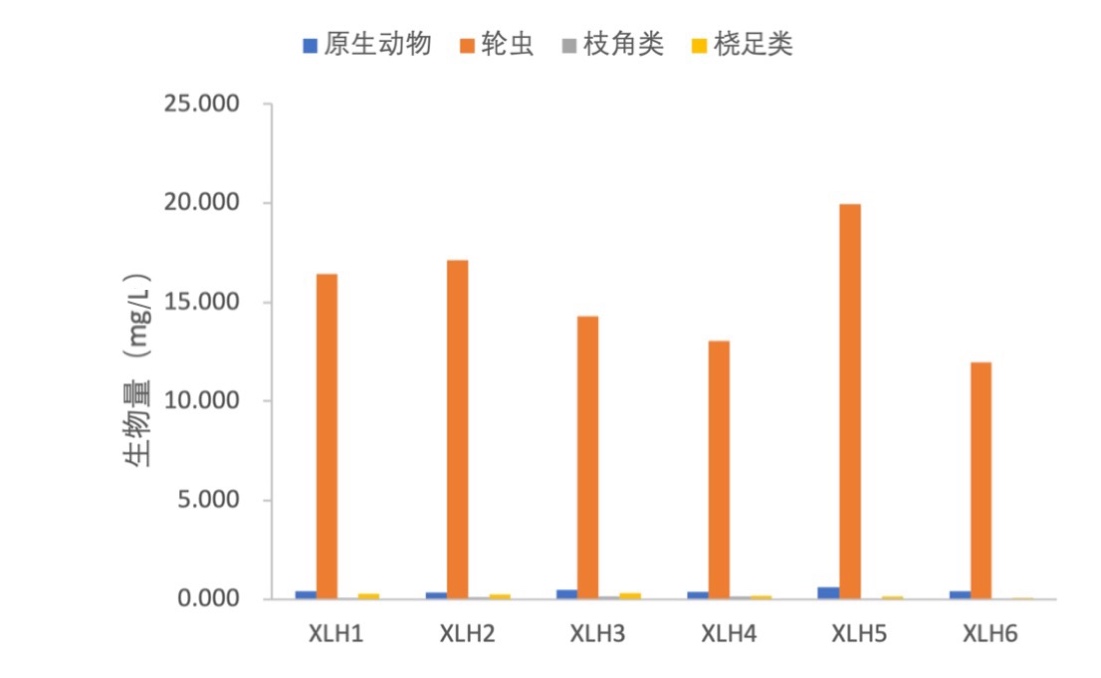
2017-2019年,原生动物密度变化范围为6890-12050 ind./L,均值为8667 ind./L;轮虫密度变化范围为9970-16640 ind./L,均值为12895 ind./L;枝角类密度变化范围为0-6 ind./L,均值为4 ind./L;桡足类密度变化范围为12-64 ind./L,均值39 ind./L(表3-15)。
原生动物生物量变化范围为0.344-0.603 mg/L,均值为0.433 mg/L;轮虫生物量变化范围为11.964-19.968 mg/L,均值为15.474 mg/L;枝角类生物量变化范围为0.009-0.118 mg/L,均值为0.075 mg/L;桡足类生物量变化范围为0.071-0.296 mg/L,均值为0.192 mg/L。XLH5浮游动物生物量最高,为20.735 mg/L;XLH6最低,为2.477 mg/L(表3-16)。浮游动物生物量与密度在各样点间差别不大,且在各样点之间的变化趋势也大致相同(图3-23)。
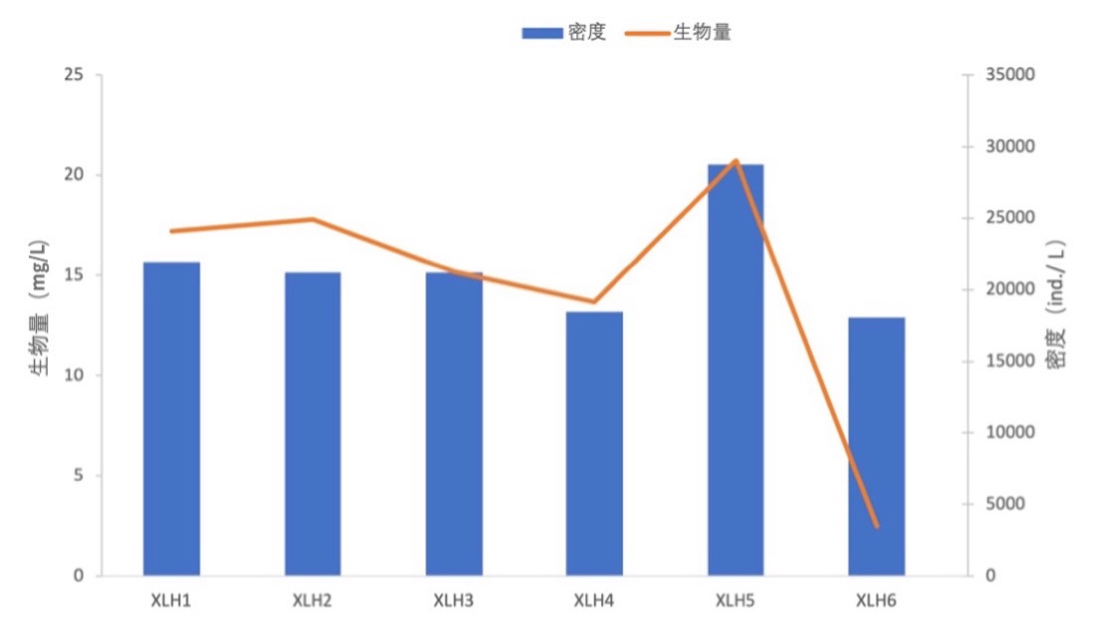
点位 | 原生动物 | 轮虫 | 枝角类 | 桡足类 | 总计 |
XLH1 | 8240 | 13700 | 4 | 64 | 21945 |
XLH2 | 6890 | 14270 | 4 | 37 | 21202 |
XLH3 | 9185 | 11930 | 6 | 64 | 21185 |
XLH4 | 7575 | 10860 | 6 | 29 | 18470 |
XLH5 | 12050 | 16640 | 2 | 27 | 28719 |
XLH6 | 8065 | 9970 | 0 | 12 | 18048 |
均值 | 8667 | 12895 | 4 | 39 | 21595 |
注:单位(ind./L)
点位 | 原生动物 | 轮虫 | 枝角类 | 桡足类 | 总计 |
XLH1 | 0.412 | 16.440 | 0.081 | 0.267 | 17.200 |
XLH2 | 0.344 | 17.124 | 0.087 | 0.228 | 17.783 |
XLH3 | 0.459 | 14.316 | 0.121 | 0.296 | 15.192 |
XLH4 | 0.379 | 13.032 | 0.118 | 0.159 | 13.688 |
XLH5 | 0.603 | 19.968 | 0.034 | 0.131 | 20.735 |
XLH6 | 0.403 | 11.964 | 0.009 | 0.071 | 2.477 |
均值 | 0.433 | 15.474 | 0.075 | 0.192 | 16.174 |
注:单位(mg/L)
2017-2019年,浮游动物优势种主要是原生动物和轮虫,原生动物和轮虫的密度占绝对优势(图3-24、3-25)。
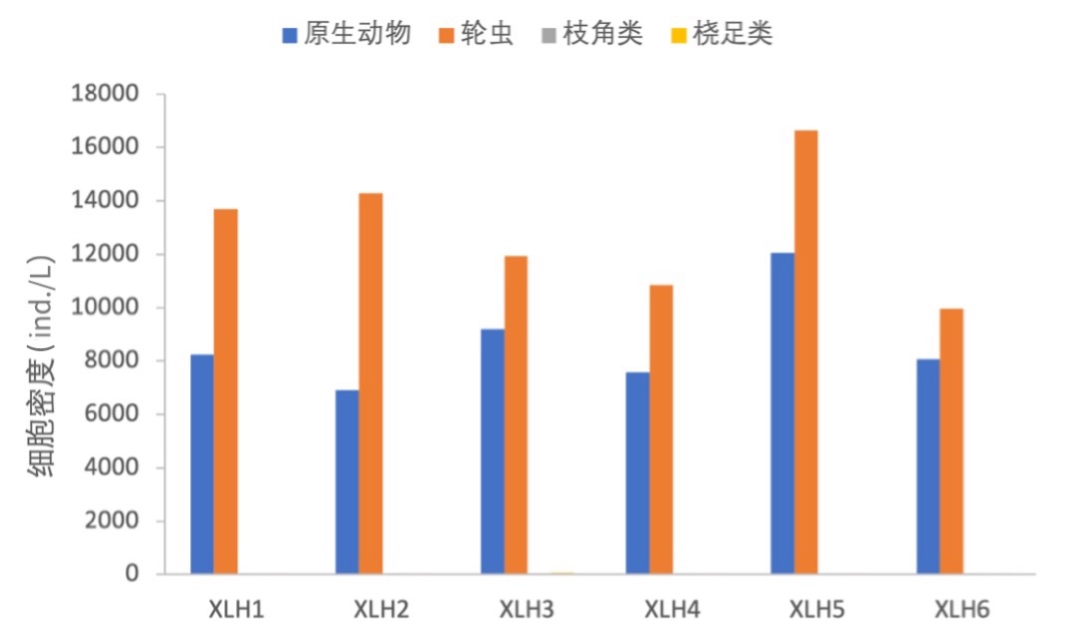
图3-25 2017-2019年西凉湖浮游动物密度组成百分比
2017-2019年,浮游动物生物量几乎以轮虫为主,其他浮游动物生物量几乎没有很大差别,轮虫生物量在全湖各样点浮游动物生物量中占据绝对优势(图3-26、3-27)。
2017-2019年,取各个采样点相对密度前十的物种进行排序,且相对密度超过10%的为优势种,浮游动物优势种主要是剑水蚤桡足幼体、无节幼体。剑水蚤桡足幼体在XLH1(32.72%)、XLH2(29.18%)、XLH6(26.31%)相对密度最大;无节幼体在XLH3(31.10%)、XLH5(26.56%)相对密度最大,XLH4相对密度最大的是王氏似铃壳虫,为27.61%(表3-17)。
种类(51种) | XLH1 | 种类(44种) | XLH2 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
剑水蚤桡足幼体 | 32.72% | + | 剑水蚤桡足幼体 | 29.18% | + |
无节幼体 | 18.06% | + | 王氏似铃壳虫 | 18.16% | + |
无棘龟甲轮虫 | 11.83% | + | 无节幼体 | 16.09% | + |
螺形龟甲轮虫 | 10.01% | + | 螺形龟甲轮虫 | 14.37% | + |
王氏似铃壳虫 | 6.63% | - | 无棘龟甲轮虫 | 6.07% | - |
脆弱象鼻溞 | 5.94% | - | 脆弱象鼻溞 | 5.47% | - |
广布多肢轮虫 | 5.01% | - | 罗氏异尾轮虫 | 4.33% | - |
圆筒异尾轮虫 | 4.71% | - | 圆筒异尾轮虫 | 2.54% | - |
罗氏异尾轮虫 | 2.70% | - | 跨立小剑水蚤 | 2.13% | - |
模糊秀体溞 | 2.40% | - | 短尾秀体溞 | 1.65% | - |
种类(46种) | XLH3 | 种类(47种) | XLH4 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
无节幼体 | 31.10% | + | 王氏似铃壳虫 | 27.61% | + |
剑水蚤桡足幼体 | 27.93% | + | 无节幼体 | 21.38% | + |
王氏似铃壳虫 | 15.43% | + | 剑水蚤桡足幼体 | 20.60% | + |
无棘龟甲轮虫 | 6.60% | - | 脆弱象鼻溞 | 10.12% | + |
螺形龟甲轮虫 | 5.17% | - | 无棘龟甲轮虫 | 6.69% | - |
脆弱象鼻溞 | 4.95% | - | 螺形龟甲轮虫 | 6.46% | - |
短尾秀体溞 | 3.08% | - | 罗氏异尾轮虫 | 2.36% | - |
罗氏异尾轮虫 | 2.39% | - | 圆筒异尾轮虫 | 1.63% | - |
模糊秀体溞 | 1.68% | - | 广布多肢轮虫 | 1.60% | - |
裂痕龟纹轮虫 | 1.68% | - | 台湾温剑水蚤 | 1.53% | - |
种类(48种) | XLH5 | 种类(45种) | XH6 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
无节幼体 | 26.56% | + | 剑水蚤桡足幼体 | 26.31% | + |
螺形龟甲轮虫 | 20.76% | + | 无节幼体 | 15.26% | + |
剑水蚤桡足幼体 | 20.01% | + | 王氏似铃壳虫 | 14.69% | + |
无棘龟甲轮虫 | 9.55% | - | 无棘龟甲轮虫 | 13.62% | + |
王氏似铃壳虫 | 7.16% | - | 螺形龟甲轮虫 | 12.55% | + |
罗氏异尾轮虫 | 4.35% | - | 圆筒异尾轮虫 | 4.42% | - |
脆弱象鼻溞 | 3.45% | - | 罗氏异尾轮虫 | 3.99% | - |
裂痕龟纹轮虫 | 3.24% | - | 广布多肢轮虫 | 3.57% | - |
广布多肢轮虫 | 3.07% | - | 哲水蚤桡足幼体 | 2.95% | - |
钟虫 | 1.83% | - | 裂痕龟纹轮虫 | 2.64% | - |
2017-2019年,共镜检出底栖动物13种,寡毛纲3种,蛭纲2种,腹足纲2种,昆虫纲6种(表3-18)。西凉湖秋季昆虫纲比例最高,为46%,其次为寡毛纲占23%,蛭纲占16%,腹足纲最少占15%(图3-28)。寡毛纲、蛭纲属于环节动物门,腹足纲属于软体动物门,昆虫纲属于节肢动物门。
种类 | 拉丁名 |
环节动物门 | Annelida |
寡毛纲 | Oligochaeta |
颤蚓科 | Tubificidae |
苏氏尾鰓蚓 | Branchiura sowerbyi |
霍甫水丝蚓 | Limnodrilus hoffmeisteri |
克拉泊水丝蚓 | Limnodrilus claparedeianus |
蛭纲 | Hirudinea |
舌蛭 | Glossiphoniidae sp. |
扁蛭 | Glossiphonia sp. |
软体动物 | Mollusca |
腹足纲 | Gastropoda |
豆螺科 | Bithyniidae |
纹沼螺 | Parafossarulus striatulus |
田螺科 | Viviparidae |
环棱螺 | Bellamya sp. |
节肢动物门 | Arthropoda |
昆虫纲 | Insecta |
蠓科 | Ceratopogonidae |
贝蠓 | Bezzia sp. |
摇蚊科 | Chironomidae |
长足摇蚊亚科 | Tanypodinae |
长足摇蚊 | Tanypus sp. |
前突摇蚊 | Procladius sp. |
直突摇蚊亚科 | Orthocladiinae |
裸须摇蚊 | Propsilocerus sp. |
德永摇蚊 | Tokunagayusurika sp. |
摇蚊亚科 | Chironominae |
摇蚊 | Chironomus sp. |
总种数 | 13 |
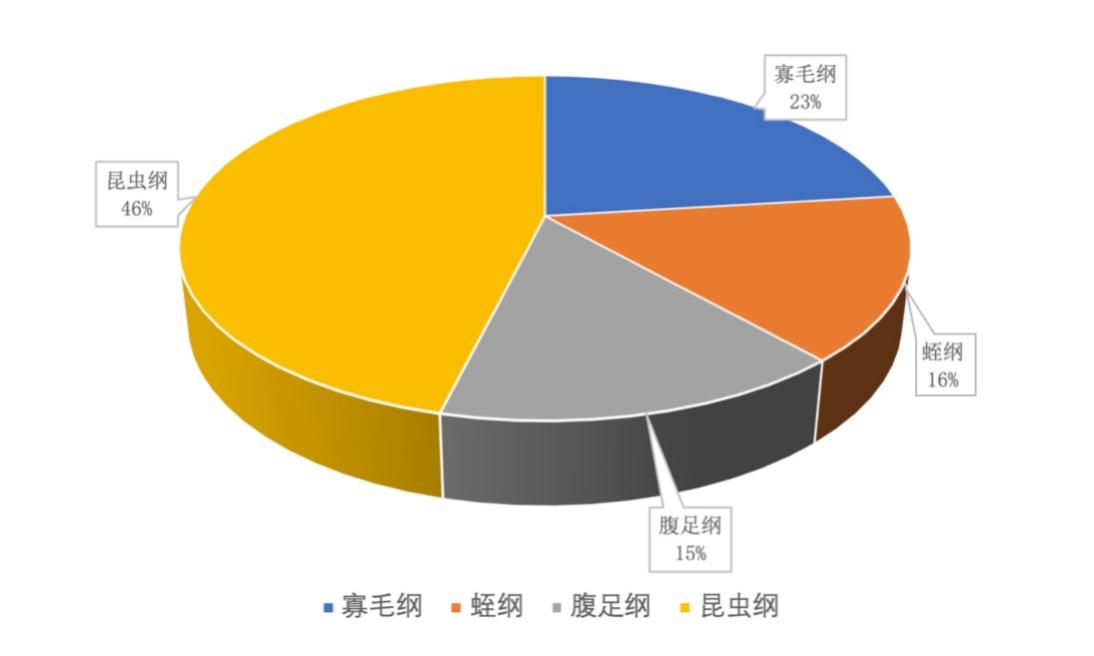
图3-28 2017-2019年西凉湖底栖动物种类组成百分比
2017-2019年,节肢动物密度变化范围为842.67-1573.33 ind/m2,均值为1239.11 ind/m2;软体动物密度变化范围为0.00-10.67 ind/m2,均值为2.67 ind/m2;环节动物密度变化范围为32.00-42.67 ind/m2,均值为34.67 ind/m2(表3-19),各样点底栖动物密度组成均为节肢动物>环节动物>软体动物。
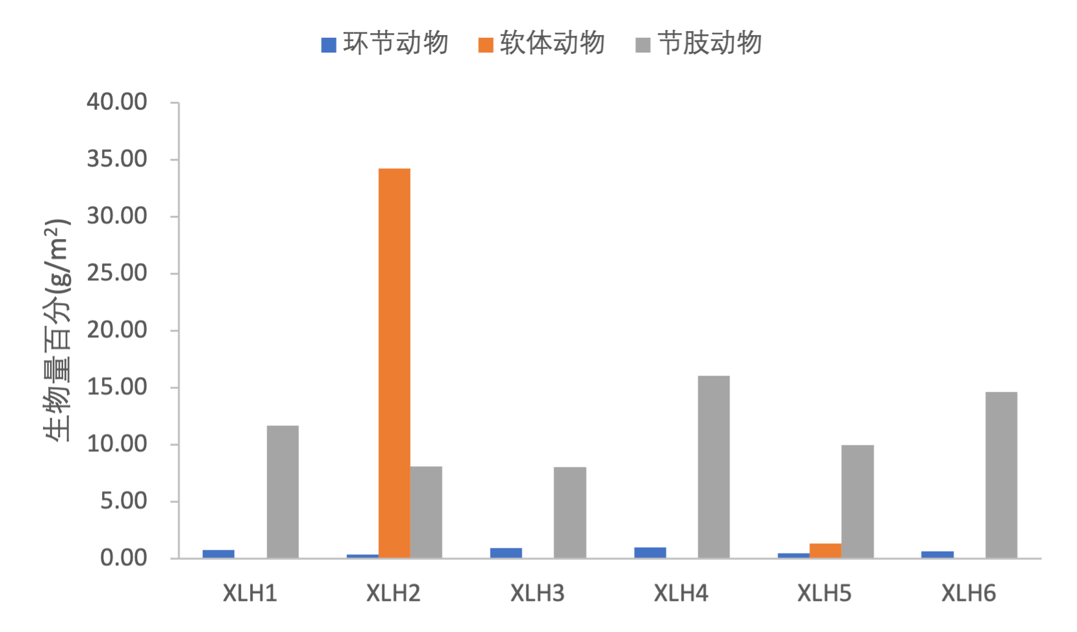
节肢动物生物量变化范围为8.07-16.04 g/m2,均值为11.42 g/m2;软体动物生物量变化范围为0.00-34.65 g/m2,均值为5.93 g/m2;环节动物生物量变化范围为0.37-1.04 g/m2,均值为0.72 g/m2(表3-20),各样点生物量组成均为节肢动物>软体动物>环节动物。软体动物只在XLH2、XLH5出现,节肢动物和环节动物各采样点均有出现。XLH2生物量最高,密度最低。(图3-29)。
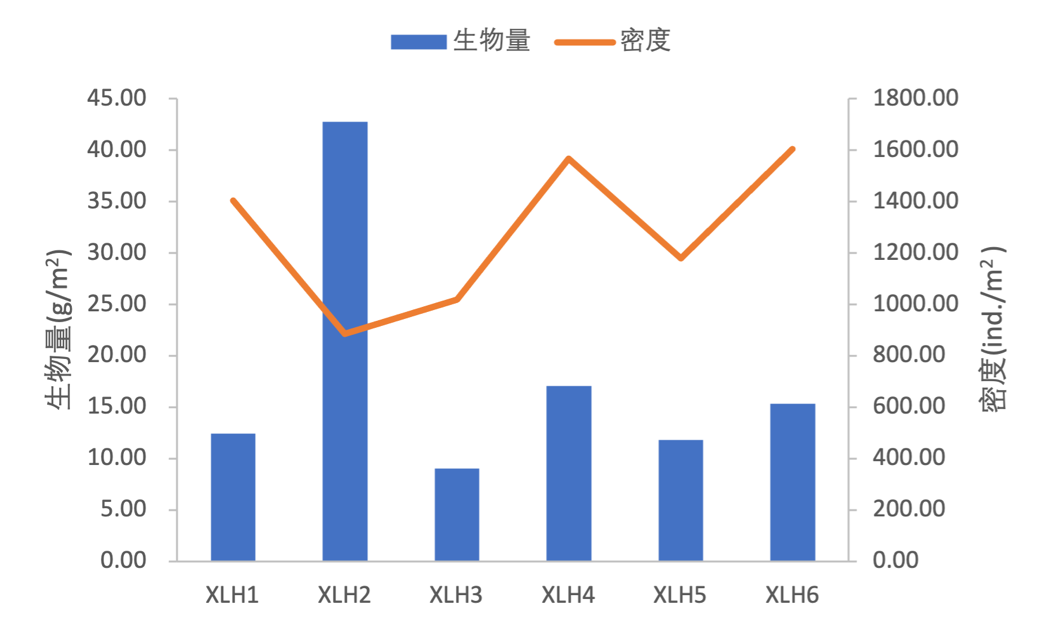
点位 | 节肢动物 | 软体动物 | 环节动物 | 总计 |
XLH1 | 1365.33 | 0.00 | 37.33 | 1402.67 |
XLH2 | 842.67 | 10.67 | 32.00 | 885.33 |
XLH3 | 976.00 | 0.00 | 42.67 | 1018.67 |
XLH4 | 1530.67 | 0.00 | 37.33 | 1568.00 |
XLH5 | 1146.67 | 5.33 | 26.67 | 1178.67 |
XLH6 | 1573.33 | 0.00 | 32.00 | 1605.33 |
均值 | 1239.11 | 2.67 | 34.67 | 1276.44 |
注:单位(ind./m2)
点位 | 节肢动物 | 软体动物 | 环节动物 | 总计 |
XLH1 | 11.67 | 0.00 | 0.77 | 12.44 |
XLH2 | 8.13 | 34.25 | 0.37 | 42.75 |
XLH3 | 8.07 | 0.00 | 0.94 | 9.01 |
XLH4 | 16.04 | 0.00 | 1.04 | 17.08 |
XLH5 | 9.96 | 1.34 | 0.51 | 11.82 |
XLH6 | 14.65 | 0.00 | 0.66 | 15.31 |
均值 | 11.42 | 5.93 | 0.72 | 18.07 |
注:单位(g/m2)
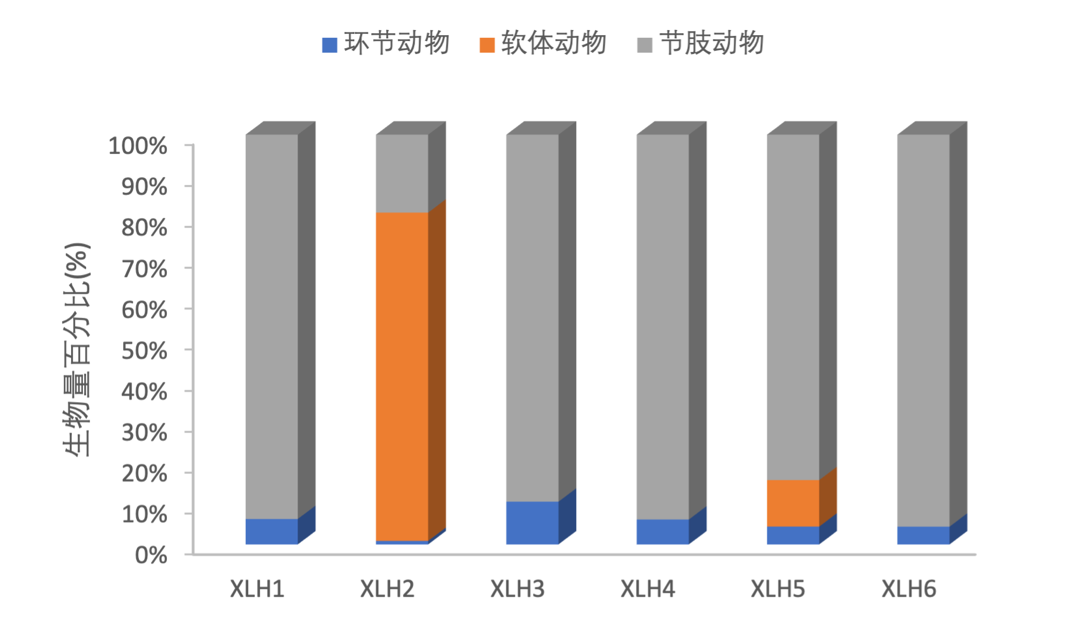
2017-2019年,底栖动物密度由环节动物、软体动物、节肢动物组成,其中节肢动物在各样点占绝对优势,软体动物和环节动物的密度在各样点均较低(图3-30、图3-31)。
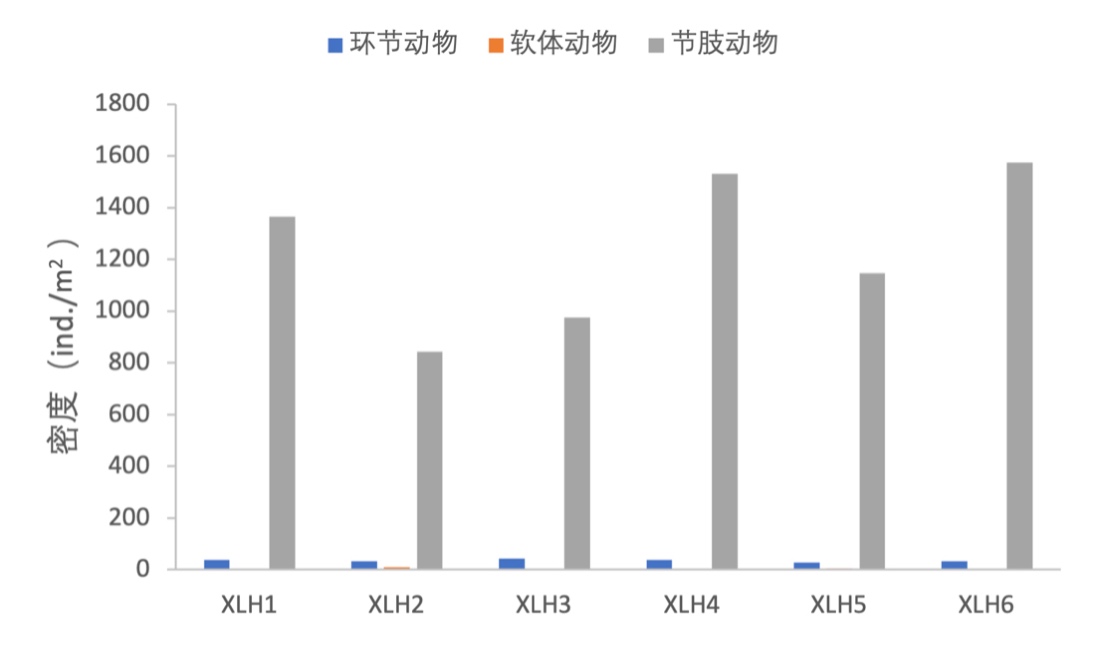
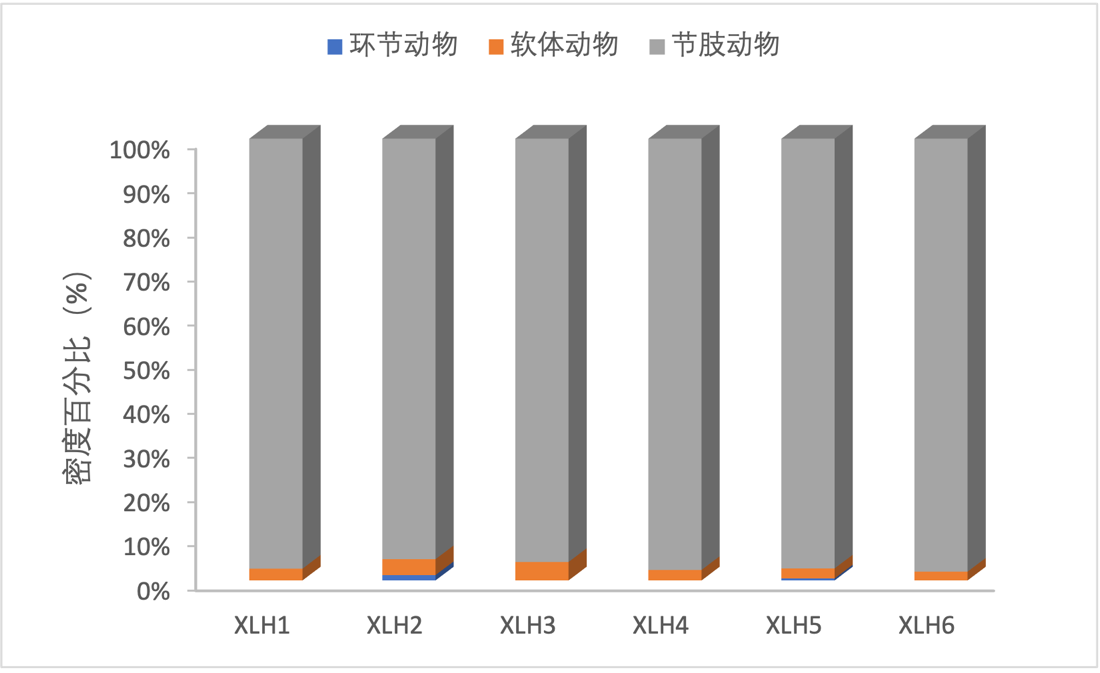
2017-2019年,底栖动物生物量组成结构主要是以节肢动物为优势种,但XLH2软体动物生物量较高,环节动物的生物量在各采样点均较低。(图3-32、图3-33)。
2017-2019年,取各个采样点相对密度前十的物种进行排序,且相对密度超过10%的为优势种。西凉湖底栖动物优势种为长足摇蚊、德永摇蚊,其中长足摇蚊为各采样点最具优势的种类,XLH2长足摇蚊相对密度最大,为66.27%(表3-21)。
种类(6种) | XLH1 | 种类(11种) | XLH2 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
长足摇蚊 | 57.79% | + | 长足摇蚊 | 66.27% | + |
德永摇蚊 | 32.32% | + | 裸须摇蚊 | 11.45% | + |
裸须摇蚊 | 5.32% | - | 德永摇蚊 | 9.64% | - |
苏氏尾鰓蚓 | 1.90% | - | 摇蚊 | 6.02% | - |
摇蚊 | 1.90% | - | 苏氏尾鰓蚓 | 1.20% | - |
扁蛭 | 0.38% | - | 舌蛭 | 1.20% | - |
环棱螺 | 1.20% | - | |||
前突摇蚊 | 1.20% | - | |||
贝蠓 | 0.60% | - | |||
霍甫水丝蚓 | 0.60% | - | |||
种类(7种) | XLH3 | 种类(7种) | XLH4 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
长足摇蚊 | 60.73% | + | 长足摇蚊 | 45.24% | + |
德永摇蚊 | 13.61% | + | 德永摇蚊 | 38.78% | + |
摇蚊 | 10.99% | + | 裸须摇蚊 | 7.14% | - |
裸须摇蚊 | 10.47% | + | 摇蚊 | 6.46% | - |
苏氏尾鰓蚓 | 2.62% | - | 苏氏尾鰓蚓 | 1.36% | - |
霍甫水丝蚓 | 1.05% | - | 霍甫水丝蚓 | 0.68% | - |
扁蛭 | 0.52% | - | 扁蛭 | 0.34% | - |
种类(8种) | XLH5 | 种类(6种) | XLH6 | ||
相对密度 | 优势种 | 相对密度 | 优势种 | ||
长足摇蚊 | 47.96% | + | 长足摇蚊 | 42.86% | + |
德永摇蚊 | 33.03% | + | 德永摇蚊 | 36.21% | + |
裸须摇蚊 | 12.22% | + | 裸须摇蚊 | 11.30% | + |
摇蚊 | 4.07% | - | 摇蚊 | 5.65% | - |
苏氏尾鰓蚓 | 1.36% | - | 苏氏尾鰓蚓 | 1.99% | - |
扁蛭 | 0.45% | - | 前突摇蚊 | 1.99% | - |
克拉泊水丝蚓 | 0.45% | - | |||
纹沼螺 | 0.45% | - | |||
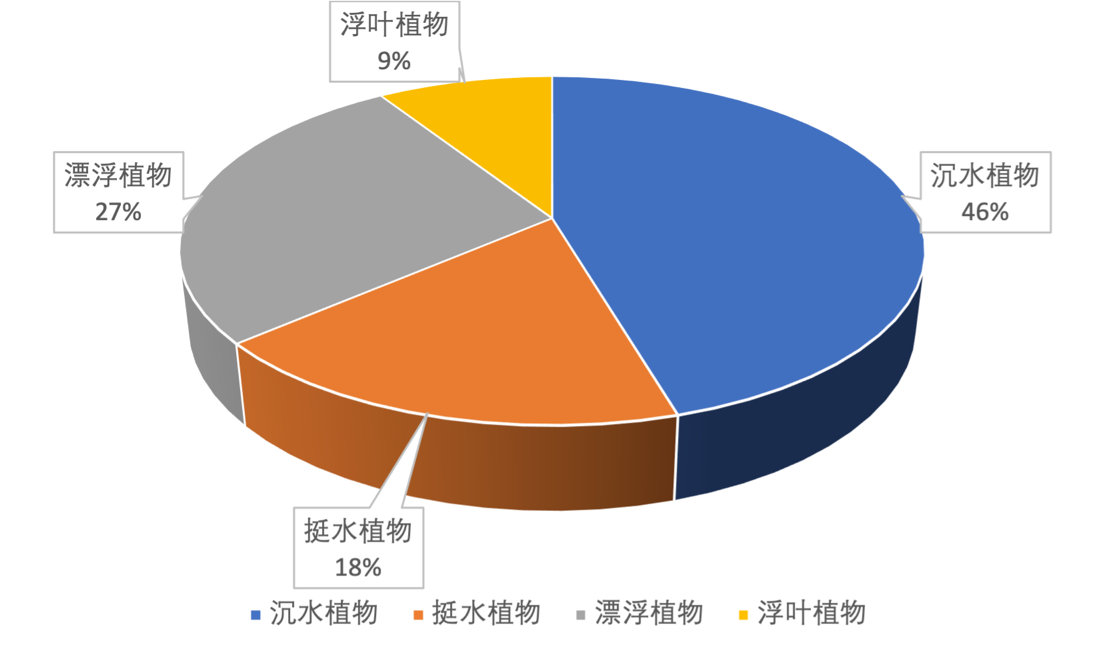
2021年4月,共鉴定出高等水生植物11种(表3-22),其中沉水植物五种:菹草、苦草、金鱼藻、穗花狐尾藻和轮叶黑藻;漂浮植物三种:凤眼莲(水葫芦)、满江红和喜旱莲子草(水花生);浮叶植物一种:菱角;挺水植物两种:菰(茭白)和芦苇(图3-34)。
种类 | 拉丁名 |
沉水植物 | |
眼子菜科 | Potamogetonaceae |
菹草 | Potamogeton crispus |
水鳖科 | Hydrocharitaceae |
苦草 | Vallisneria natans |
轮叶黑藻 | Hydrilla verticillata |
金鱼藻科 | Ceratophyllaceae |
金鱼藻 | Ceratophyllum demersum |
小二仙草科 | Haloragidaceae |
穗花狐尾藻 | Myriophyllum spicatum |
漂浮植物 | |
苋科 | Amaranthaceae |
喜旱莲子草 | Alternanthera sessilis |
雨久花科 | Potederiaceae |
凤眼莲 | Eichhornia crassipes |
满江红科 | Azollaceae |
满江红 | Azolla imbricata |
浮叶植物 | |
菱科 | Trapaceae |
菱角 | Trapa japonica |
挺水植物 | |
禾本科 | Gramineae |
菰 | Zizania caduciflora |
芦苇 | Phragmites communis |
总计 | 11 |
2020-2021年,沉水植物生物量平均为163.5 g/m2,在沉水植物分布方面,东湖区的沉水植物分布面积和生物量均大于西湖区,具体表现为东湖区全湖分布较为均匀,而西湖区中间湖区部分沉水植物很少,大部分集中在西南角和东北角湖边(图3-35)。
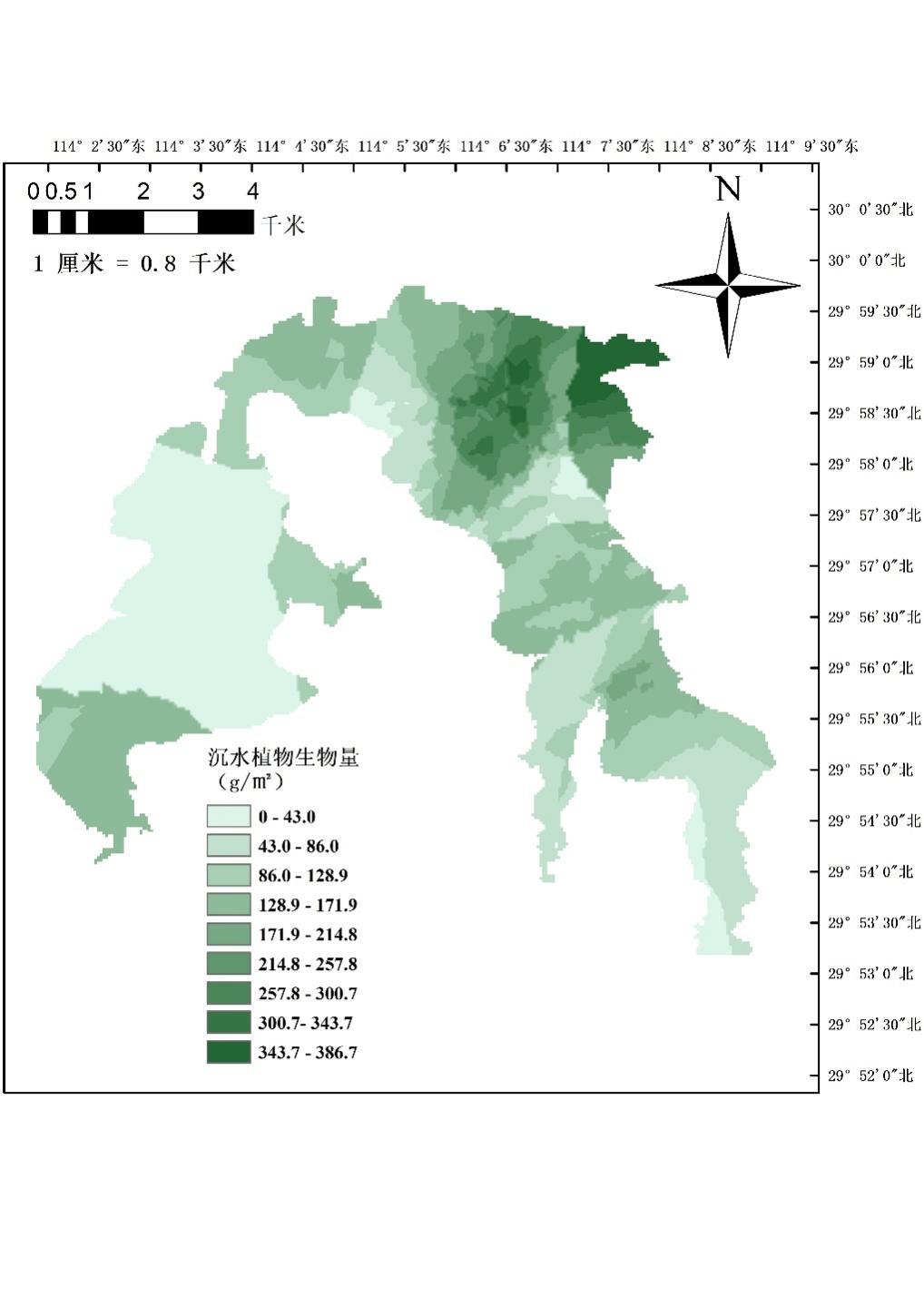
沉水植物优势种为菹草和苦草,其中轮叶黑藻和苦草只在东湖区采集到样本,金鱼藻和穗花狐尾藻分布区域较少,虽然收集到该种的标本,但是并未定量采集到该种类(图3-36 、表3-23)。
湖区 | 菹草 | 苦草 | 轮叶黑藻 |
西湖区 | 83.16 | 0 | 0 |
东湖区 | 94.443 | 44.49 | 0.15 |
注:单位(g/m2)
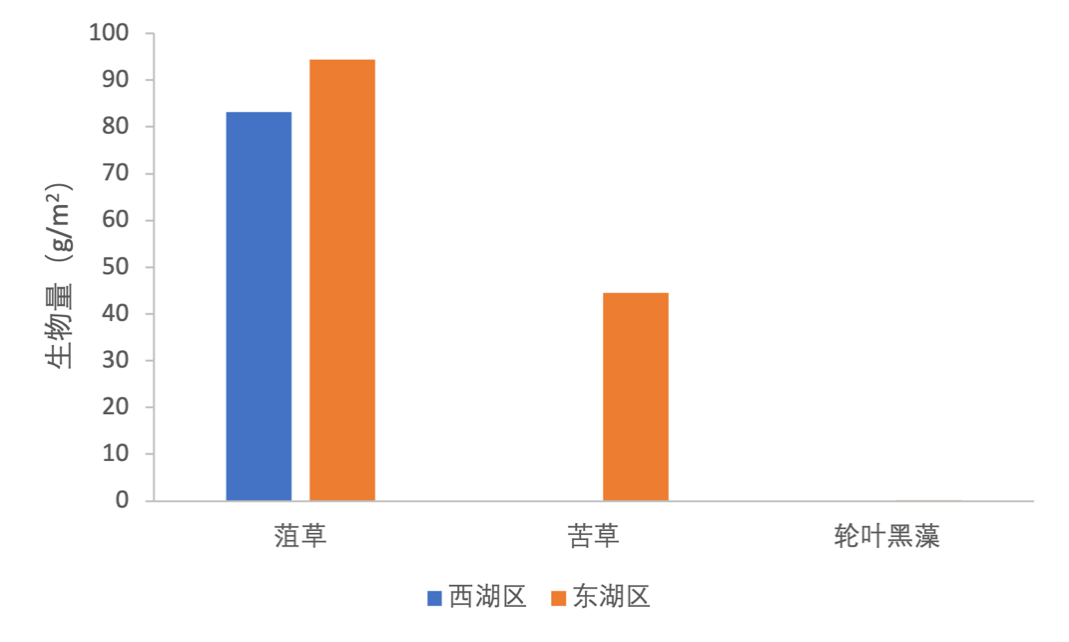
2020-2021年,共鉴定鱼类25种,共计7目6科(表3-24)。鲤科鱼类种类最多,占总物种数的80%,其中鲌亚科在鲤科内种类数最多,其他科种类数相同,均为总物种数的4%(图3-37)。
种类 | 拉丁名 | 2020年11月 | 2021年04月 |
鲱形目 | Clupeiformes | ||
鳀科 | Engraulidae | ||
湖鲚 | Coilia nasus | √ | √ |
鲤形目 | Cypriniformes | ||
鲤科 | Cyprinidae | ||
鲃亚科 | Barbinae | ||
麦瑞加拉鲮 | Cirrhinus mrigala | √ | √ |
雅罗鱼亚科 | Leuciscinae | ||
草鱼 | Ctenopharyngodon idellus | √ | √ |
鲌亚科 | Culterinae | ||
贝氏䱗 | Hemiculter bleekeri | √ | √ |
䱗 | Hemiculter leucisculus | √ | √ |
似鱎 | Toxabramis swinhonis | √ | √ |
红鳍原鲌 | Cultrichthys erythropterus | √ | √ |
翘嘴鲌 | Culter alburnus | √ | √ |
达氏鲌 | Culter dabryi | √ | √ |
团头鲂 | Megalobrama amblycephala | √ | √ |
鲴亚科 | Xenocypriniae | ||
湖北圆吻鲴 | Distoechodon hupeinensis | √ | |
似鳊 | Pseudobrama simoni | √ | √ |
鲢亚科 | Hypophthalmichthyinae | ||
鳙 | Aristichthy nobilis | √ | |
鲢 | Hypophthalmichthys molitrix | √ | √ |
鮈亚科 | Gobioninae | ||
麦穗鱼 | Pseudorasbora parva | √ | √ |
银鮈 | Squalidus argentatus | √ | |
棒花鱼 | Abbottina rivularis | √ | |
鱊亚科 | Acheilognathinae | ||
大鳍鱊 | Acheilognathus macropterus | √ | √ |
兴凯鱊 | Acheilognathus chankaensis | √ | √ |
鲤亚科 | Cyprininae | ||
鲤 | Cyprinus carpio | √ | √ |
鲫 | Carassius auratus | √ | √ |
鲇形目 | Siluriformes | ||
鮰科 斑点叉尾鮰 | Ictaluridae Ictalurus Punetaus | √ | |
颌针鱼目 | Beloniformes | ||
鱵科 | Hemiramphidae | ||
间下鱵 | Hyporhamphus intermedius | √ | |
鲈形目 | Perciformes | ||
鰕虎鱼科 | Gobiidae | ||
子陵吻虾虎鱼 | Rhinogobius giurinus | √ | |
合鳃目 | Symbranchiformes | √ | √ |
合鳃科 | Symbranchidae | ||
黄鳝 | Monopterus albus | √ | |
总计 | 22 | 22 | |
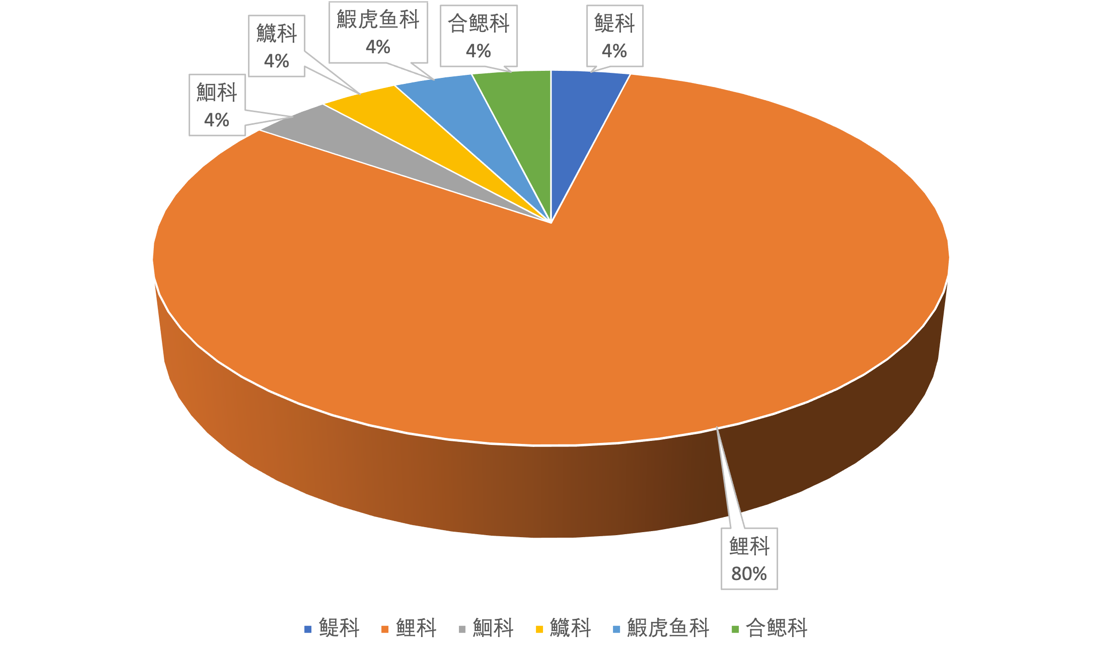
2020-2021年,鱼类群落调查分东湖区(6个样点)、西湖区(6个样点)和中间连通水道(3个样点)三个区域进行,每个样点各下一组网具(一条沉性复合刺网,一条浮性复合刺网,一条地笼),鱼类密度和生物量分别用单位努力捕捞密度(NPUE,ind. net-1 h-1)和单位努力捕捞生物量(BPUE,g net-1 h-1)来表示,每组网具单位努力捕捞量计算方法为统计每小时捕获的鱼类数量和重量。
根据调查显示,东、西湖区鱼类密度相差不大,东湖区XLH15鱼类密度最大,XLH9最低。鱼类生物量表现为西湖区>水道>东湖区,XLH1鱼类生物量最大。东湖区和西湖区鱼类密度和生物量各样点之间的趋势大致相同(图3-38、图3-39)。
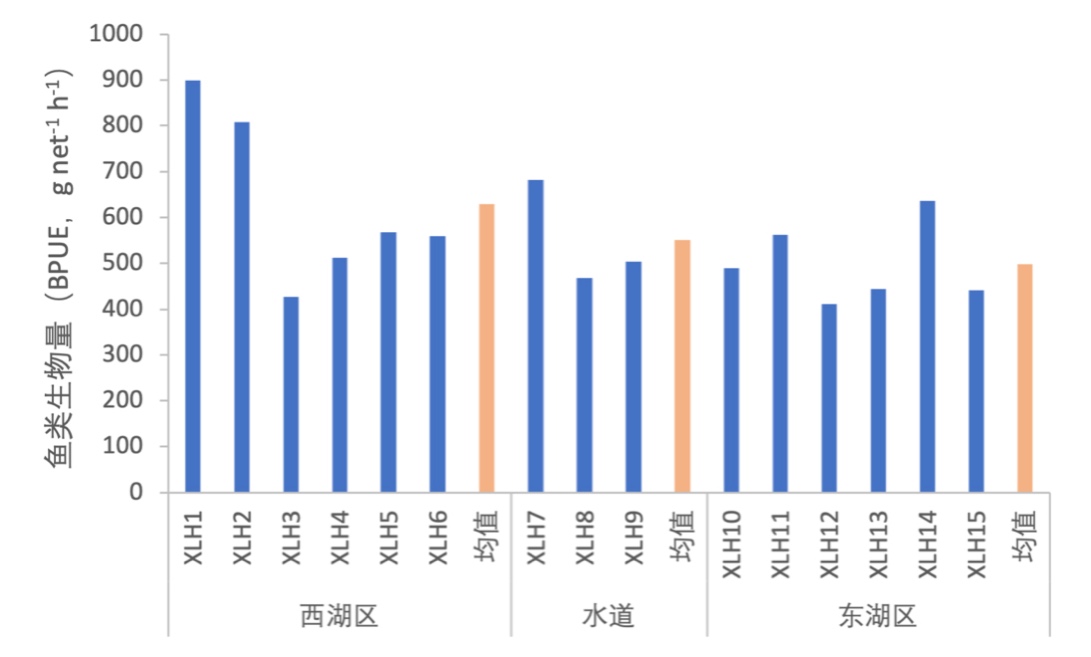
3.7.3.1 活动水层划分
根据鱼类生活水层分布情况,将鱼类分为中上层鱼类、中下层鱼类和底层鱼类三类。按照鱼类密度统计,中上层鱼类占绝对优势,其中西湖区中上层鱼类较东湖区更多(图3-40);按照鱼类生物量统计,中上层鱼类生物量仍为最高,但中下层鱼类比例更接近中上层鱼类,这证明中下层鱼类个体较大;无论按照密度和生物量统计,底层鱼类占比均很小(图3-41)。
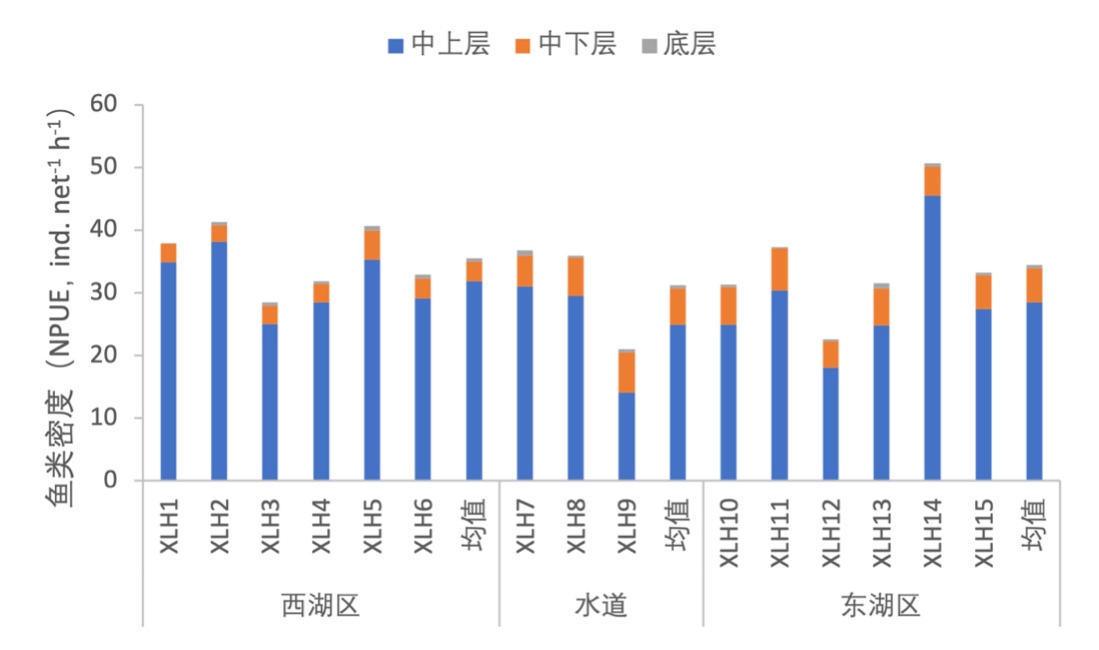
图3-40 2020-2021年西凉湖鱼类活动水层组成(密度)
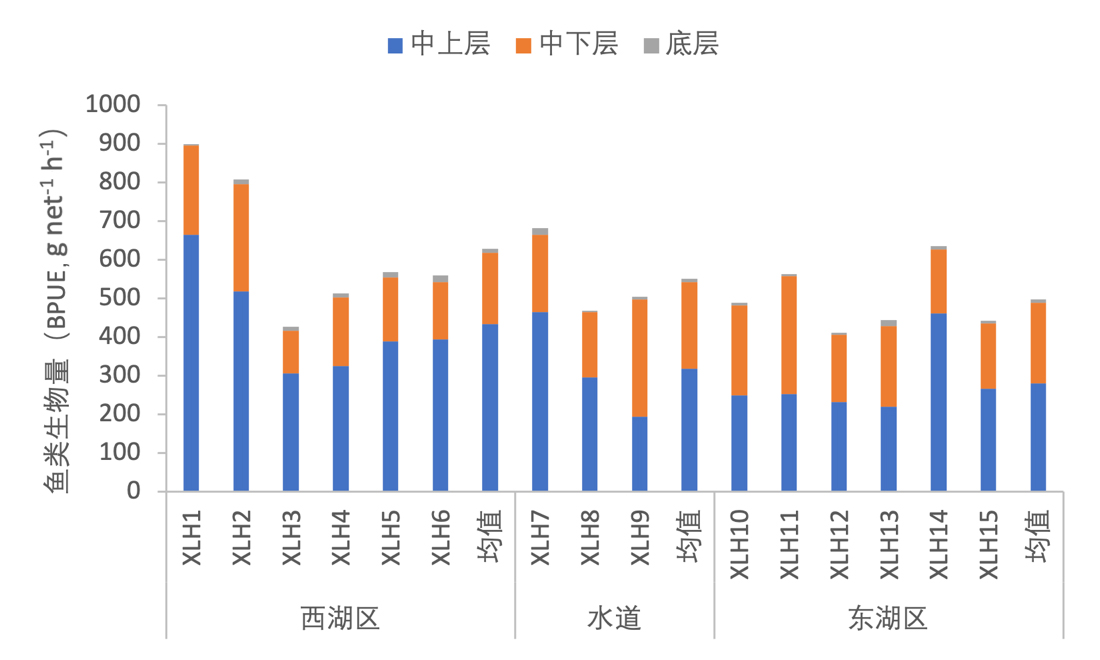
图3-41 2020-2021年西凉湖鱼类活动水层分布(生物量)
3.7.3.2 摄食习性划分
根据鱼类摄食习性的不同,将鱼类食性划分为草食性、浮游动物食性、底栖动物食性、食鱼性、碎屑食性、杂食性和藻类食性。不同的鱼类对于食物的选择不同,同一种食性的鱼类存在竞争现象,鱼类群落食性的单一化,往往会影响鱼类群落的多样性。2020-2021年,根据西凉湖鱼类密度以及生物量分布情况,对鱼类食性结构进行分析。按照鱼类密度对食性组成进行分析,以浮游动物食性占绝对优势,杂食性鱼类也具有一定的密度,其他食性相对较少(图3-42)。按照鱼类生物量对食性组成进行分析,以浮游动物食性占主要优势,杂食性和藻类食性的鱼类也占有较高的比例(图3-43)。综上所述,浮游动物食性鱼类体型较小,并且在数量上占有绝对优势;藻类食性和杂食性鱼类体型较大。
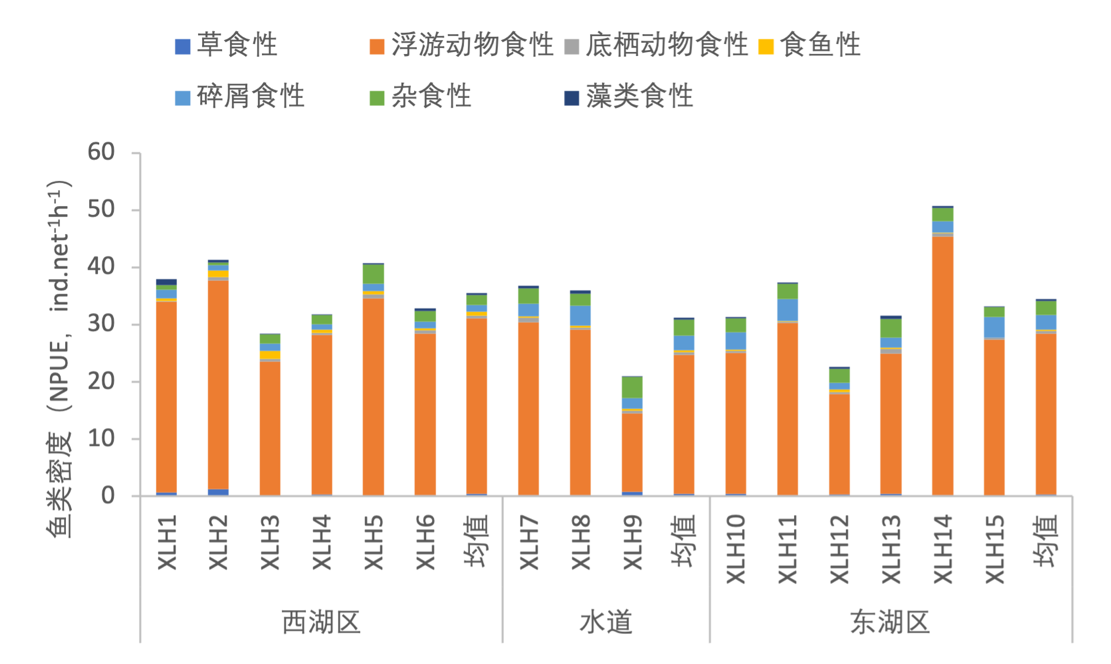
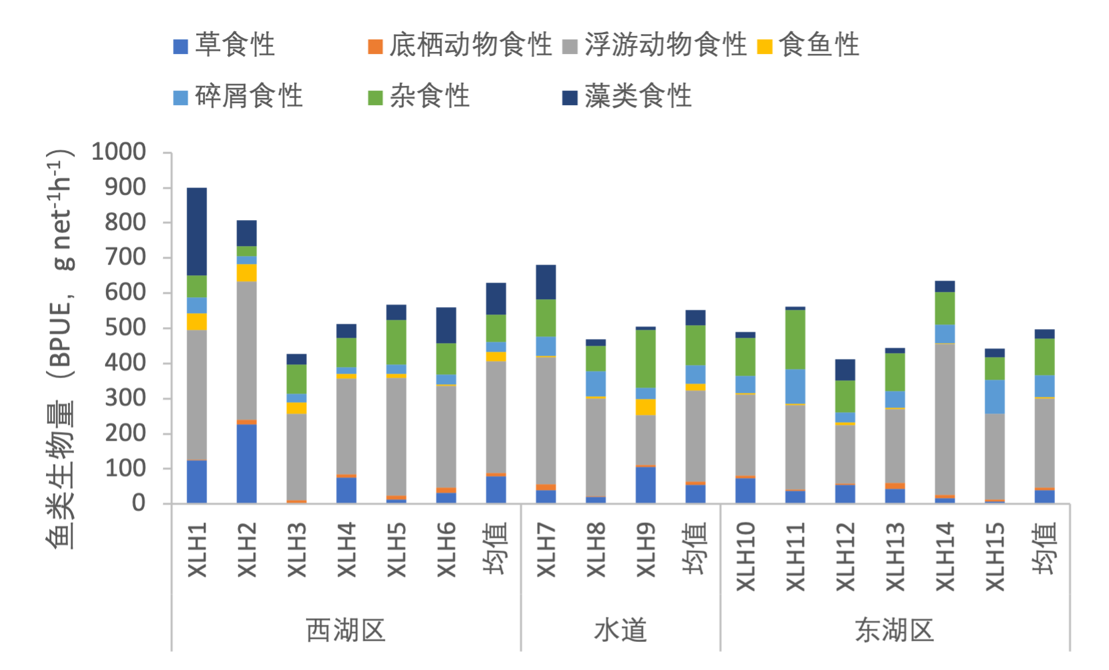
图3-43 2020-2021年西凉湖鱼类食性组成(生物量)
3.7.3.3 繁殖习性划分
根据鱼类繁殖行为的不同,将鱼类繁殖习性划分为广适型、静水型、江河洄游型和流水型。不同的繁殖习性会导致鱼类产卵场不同。2020-2021年,西凉湖鱼类群落主要是以静水型为主,按照鱼类密度对繁殖习性组成进行分析,静水型鱼类远大于其他类型的鱼类(图3-44);按照鱼类生物量对繁殖习性组成进行分析,除静水型鱼类占据较大比例外,江河洄游型鱼类也占有一定比例,并且西湖区江河洄游型鱼类较东湖区具有较大的生物量(图3-45)。
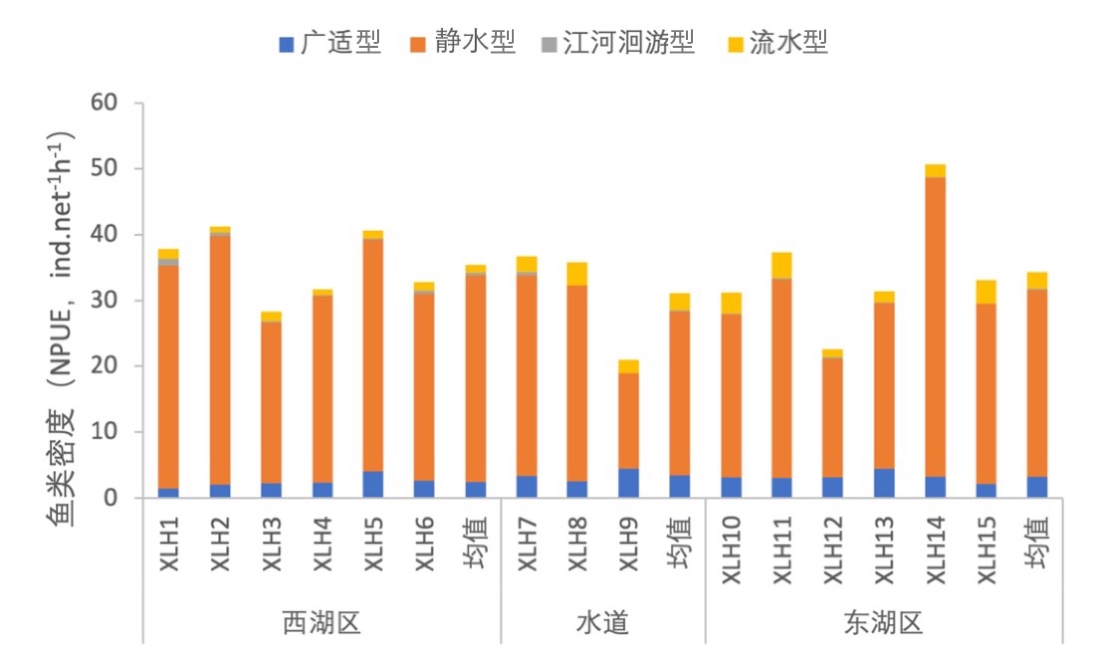
图3-44 2020-2021年西凉湖鱼类繁殖习性组成(密度)
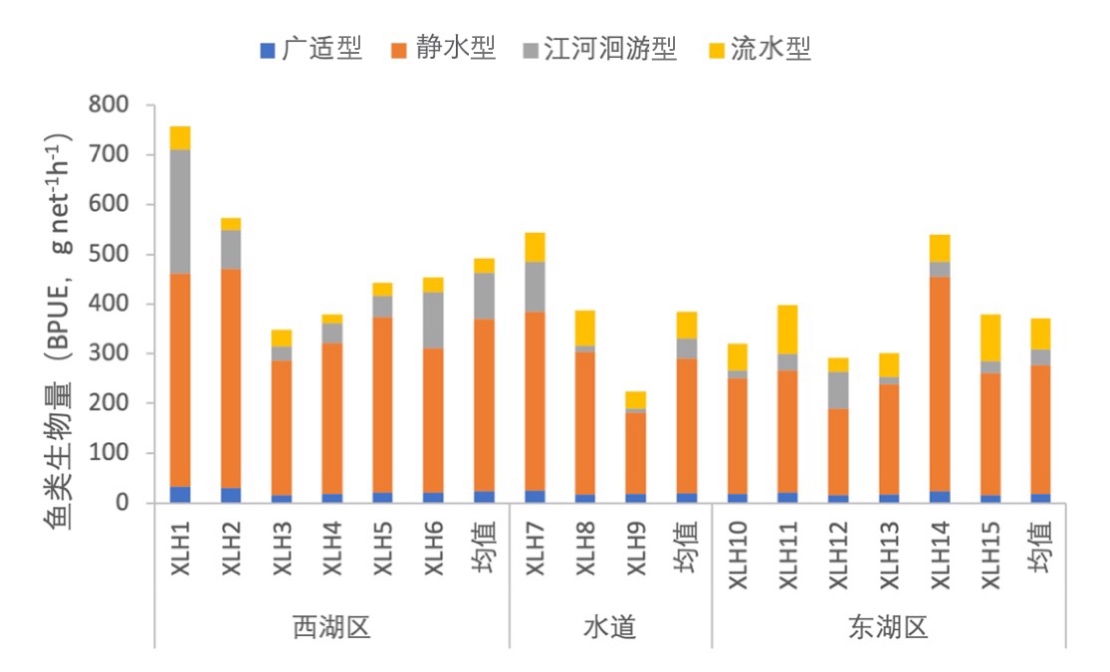
图3-45 2020-2021年西凉湖鱼类繁殖习性组成(生物量)
3.7.3.4 鱼卵生态类型划分
根据鱼卵性状的不同,将鱼类与卵的生态类型划分为沉性、浮性、漂流性和黏性,一些鱼类产卵必须要在特定的条件下才能进行,需要外部环境条件的刺激,这代表着该类鱼类需要较为苛刻的产卵条件。2020-2021年,按照鱼类密度对产不同生态类型卵的鱼类组成进行分析,产粘性卵的鱼类几乎占据整个西凉湖区(图3-46);按照鱼类生物量进行分析,产粘性卵的鱼类占绝对优势,但产漂流性卵的鱼类也占有一定的比例,并且产漂流性卵的鱼类在西湖区具有较高的生物量(图3-47)。
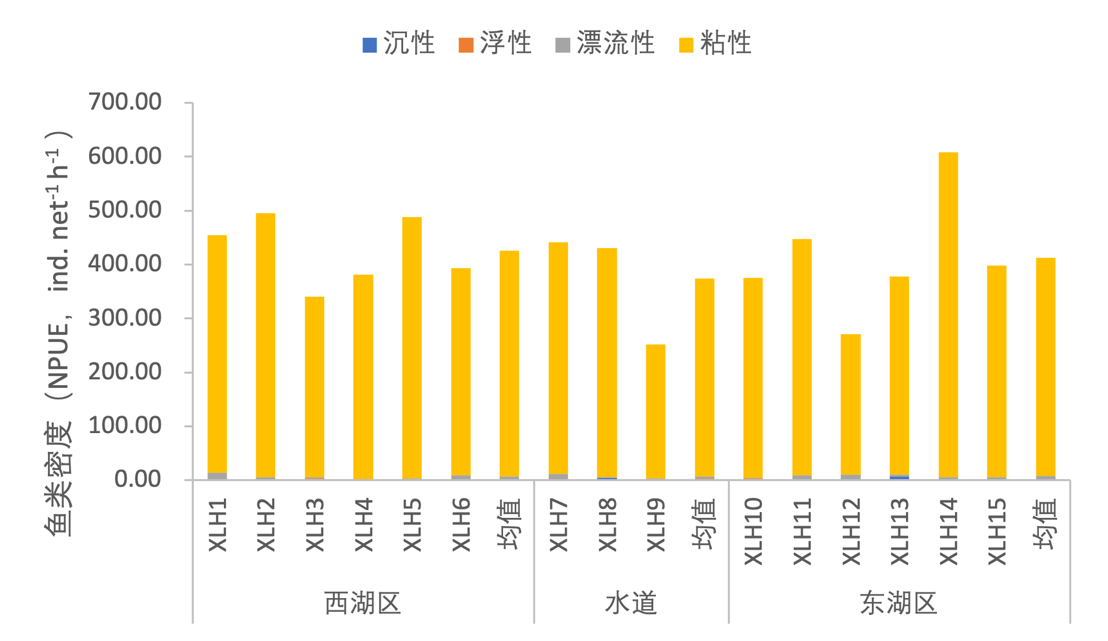
图3-46 2020-2021年西凉湖鱼类受精卵类型组成(密度)
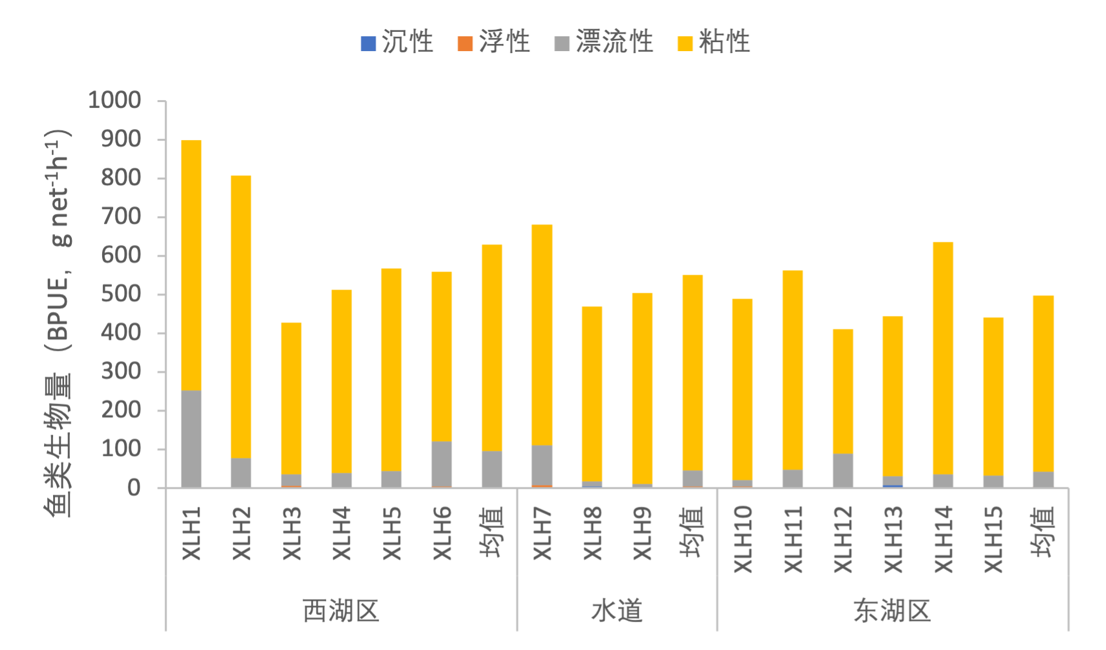
图3-47 2020-2021年西凉湖鱼类受精卵类型组成(生物量)
本研究采用水生生物群落常用于优势种确定的Pinkas相对重要性指数(Index of Relative Importance,IRI),计算公式为:
IRI = ( N% + B% )×F%×10000
式中:N%为某一种类的数量占总数百分比,B%为某一种类的重量占总重百分比,F%为某一种类在捕捞中出现的次数占总捕捞次数的百分比。Pinkas相对重要性指数对各组湖泊鱼类的重要性进行分类:IRI > 1000的种类为优势种,100 < IRI < 1000的种类为重要种,10 < IRI < 100的种类为常见种,1 < IRI < 10的种类为一般种,IRI < 1的种类为少见种。
2020-2021年西凉湖鱼类组成中,优势种为似鱎;重要种为:䱗、红鳍原鲌、鲫;常见种为:达氏鲌、黄颡鱼、鲢、麦瑞加拉鲮;一般种为草鱼、大鳍鱊、鲤、麦穗鱼、似鳊;少见种为:斑点叉尾鮰、棒花鱼、贝氏䱗、短颌鲚、湖北圆吻鲴、间下鱵、翘嘴鲌、团头鲂、兴凯鱊、银鮈、鳙、子陵吻虾虎鱼、黄鳝。其中䱗、似鱎、鲫、麦瑞加拉鲮在每个采样点均有出现,西凉湖鱼类优势种以小型鱼类为主,大型鱼类和凶猛型鱼类优势度较低(表3-25)。
种类 | IRI指数 | 重要性分类 | 种类 | IRI指数 | 重要性分类 |
似鱎 | 1121.0737 | 优势种 | 团头鲂 | 0.9421 | 少见种 |
鲫 | 211.6901 | 重要种 | 贝氏䱗 | 0.8294 | 少见种 |
䱗 | 202.5288 | 重要种 | 子陵吻虾虎鱼 | 0.7929 | 少见种 |
红鳍原鲌 | 129.5876 | 重要种 | 翘嘴鲌 | 0.4690 | 少见种 |
鲢 | 57.7750 | 常见种 | 湖北圆吻鲴 | 0.2577 | 少见种 |
麦瑞加拉鲮 | 49.3556 | 常见种 | 间下鱵 | 0.2223 | 少见种 |
达氏鲌 | 27.2522 | 常见种 | 兴凯鱊 | 0.0997 | 少见种 |
黄颡鱼 | 25.1280 | 常见种 | 短颌鲚 | 0.0568 | 少见种 |
似鳊 | 4.8444 | 一般种 | 鳙 | 0.0181 | 少见种 |
麦穗鱼 | 2.7358 | 一般种 | 斑点叉尾鮰 | 0.0120 | 少见种 |
大鳍鱊 | 1.7662 | 一般种 | 黄鳝 | 0.0095 | 少见种 |
鲤 | 1.1601 | 一般种 | 银鮈 | 0.0067 | 少见种 |
草鱼 | 1.0445 | 一般种 | 棒花鱼 | 0.0033 | 少见种 |
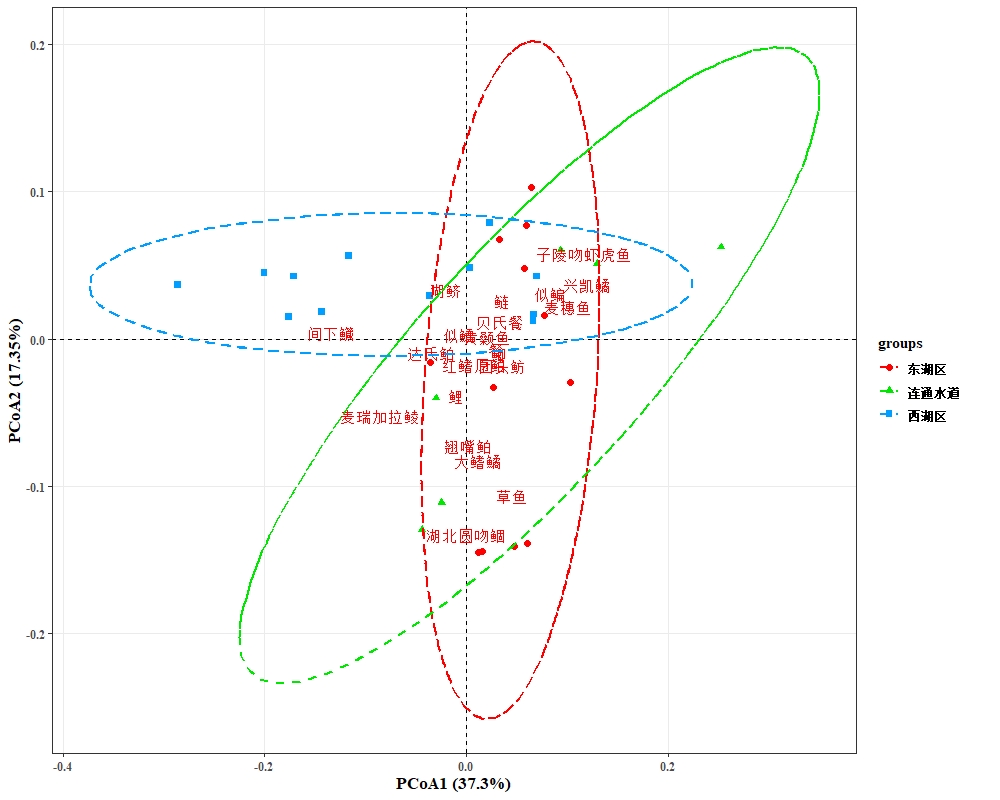
根据鱼类群落分布特征,对西凉湖2020-2021年鱼类群落进行主坐标分析(PCoA),西湖区与东湖区具有显著性差异(P<0.05),连通水道与西湖区没有显著性差异(P>0.05), 连通水道与东湖区没有显著性差异(P>0.05)。东湖区中大鳍鱊、草鱼、翘嘴鲌、湖北圆吻鲴相对较多;西湖区中间下鱵、鲢、贝氏䱗相对较多。东湖区水草较多,这可能是大鳍鱊、草鱼等食草性鱼类多的原因,而翘嘴鲌属于肉食性鱼类,可能是由于东湖区小型鱼类较多提供了丰富的食物;西湖区浮游生物含量丰富,所以滤食性和浮游生物食性的鱼类较多(图3-48)。
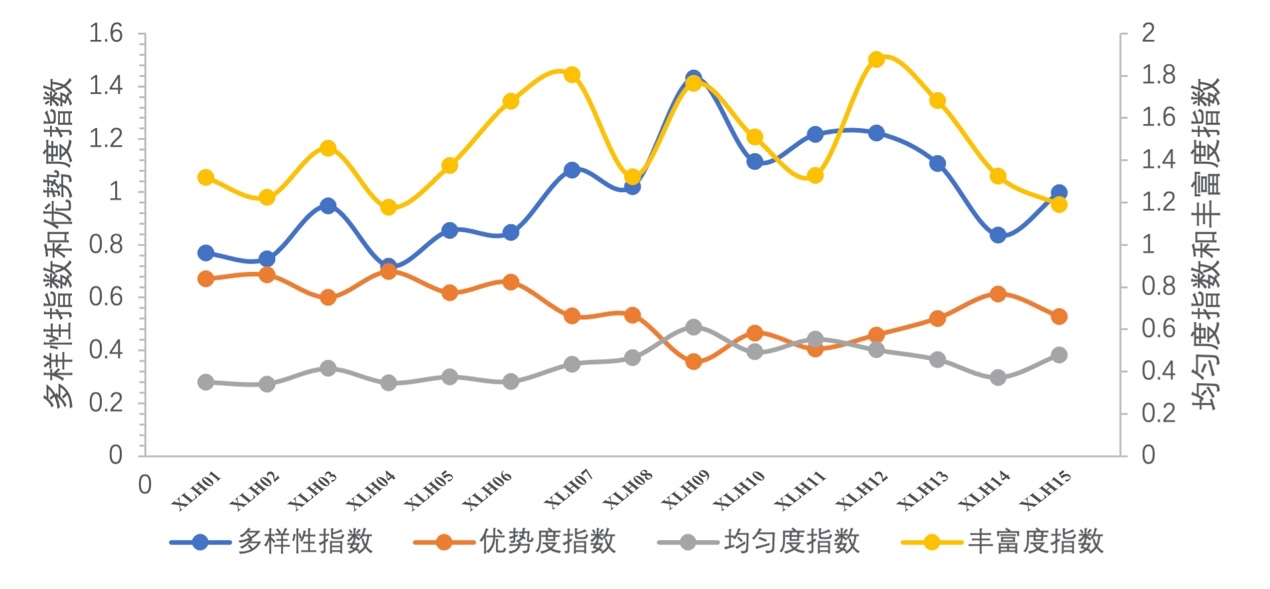
对西凉湖2020-2021鱼类群落生物多样性进行分析,均匀度指数变化比较均匀;丰富度指数变化变化较大,其中XLH7、9、12最高;优势度指数总体西湖区略高于东湖区;多样性指数总体东湖区略高于西湖区;XLH9丰富度和多样性均最高(图3-49)。
3.8.1.1 浮游生物现存量
按照《大水面生态增养殖容量计算方法(SC/T 1149-2020)》,根据2017-2019年西凉湖浮游植物、浮游动物的本底调查资料,西凉湖水域面积85.20 km2,平均水深2 m,计算出浮游植物现存量2559.4 t,浮游动物为2755.3 t(表3-26)。
生物类群 | 生物量(mg/L) | 正常库容(m3) | 现存量(t) |
浮游植物 | 15.02 | 1.704×108 | 2559.4 |
浮游动物 | 16.17 | 1.704×108 | 2755.3 |
3.8.1.2 底栖动物现存量
按照《大水面生态增养殖容量计算方法(SC/T 1149-2020)》,根据2017-2019年西凉湖底栖动物的本底调查资料,西凉湖水域面积85.20 km2,计算出寡毛类现存量为56.40 t,蛭类为6.73 t,腹足类为758.11 t,水生昆虫为972.98 t(表3-27)。
生物类群 | 生物量(g/m2) | 面积(km2) | 现存量(t) |
寡毛类 | 0.662 | 85.20 | 56.40 |
蛭类 | 0.079 | 85.20 | 6.73 |
腹足类 | 8.898 | 85.20 | 758.11 |
水生昆虫 | 11.420 | 85.20 | 972.98 |
3.8.1.3 沉水植物现存量
西凉湖高等水生植物的现存量为:菹草4.93×104 t,苦草1.87×104 t,轮叶黑藻639.0 t(表3-28)。
种类 | 生物量(g/m2) | 面积(km2) | 现存量(t) |
菹草 | 578.50 | 85.20 | 4.93×104 |
苦草 | 219.50 | 85.20 | 1.87×104 |
轮叶黑藻 | 7.50 | 85.20 | 639.0 |
3.8.2.1 浮游生物渔业潜力评估
采用生物量转化法,根据浮游生物现存量生物量,利用率,P/B系数以及饵料系数来估算鱼产力,一般计算公式为:
鱼产力=浮游生物现存量生物量*利用率*P/B系数/饵料系数
根据相关文献中记载,不同地区、不同环境以及不同气候等原因会导致鱼产力。参考《大水面增养殖容量计算方法》中华中地区取值,浮游植物的P/B系数取值为100,浮游动物的取值为30。同时按照《大水面增养殖容量计算方法》规定,浮游植物饵料系数取值为100,最大利用率取值为40%;浮游动物饵料系数取值为10,最大利用率取值为50%。
根据2017-2019年调查的数据,西凉湖浮游植物和浮游动物的鱼产力分别估算为1663.61 t/年和2479.77 t/年,因此西凉湖浮游生物的鱼产力总计为4143.38 t/年(表3-29)。
生物类群 | 现存量(t) | 利用率(%) | P/B系数 | 饵料系数 | 鱼产力(t) |
浮游植物 | 2559.4 | 40 | 130 | 80 | 1663.61 |
浮游动物 | 2755.3 | 30 | 30 | 10 | 2479.77 |
总计 | 4143.38 | ||||
采用能量估算法,求出底栖动物对鱼类的供饵能力,进而计算其鱼产力。由于水生昆虫、水栖寡毛类和软体动物的生物特性(如P/B系数、干湿重比、能值等)不尽相同,其计算公式略有差别。按梁彦龄等(1995)给出的公式进行估算:
水生昆虫的鱼产力估算:FI = 0.183 BI
水栖寡毛类的鱼产力估算:Fo = 0.235 Bo
软体动物的鱼产力估算:FM = 0.032 BM
根据2017-2019年调查数据西凉湖寡毛类鱼产力估算为13.25 t/年,腹足类为25.26 t/年,水生昆虫类为178.06 t/年,总计216.57 t/年(表3-30)。
生物类群 | 现存量 | 转换系数 | 鱼产力(t) |
寡毛类 | 56.40 | 0.235 | 13.25 |
腹足类 | 758.11 | 0.032 | 25.26 |
水生昆虫类 | 972.98 | 0.183 | 178.06 |
总计 | 216.57 | ||
3.8.2.3 高等水生植物渔业潜力评估
采用能量估算法,求出高等植物水生植物对鱼类的供饵能力(表3-31)。进而计算出鱼产力。水生维管束植物的利用率为25%,饵料系数为100,计算公式为:
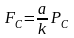
FC——草食性鱼类或其它水生经济动物的增殖容量,单位为吨(t)
PC——水生维管束植物年净生产量,单位为吨(t)
a——鱼类等水生经济动物对该类饵料生物允许的最大利用率;不同生态类型鱼类等水生经济动物对不同饵料生物允许的最大利用率
k——鱼类等水生经济动物对该类饵料生物的饵料系数,不同生态类型鱼类等水生经济动物对不同饵料生物的饵料系数
表3-31 2017-2019年西凉湖高等水生植物鱼产力估算
生物类群 | 现存量 | 利用率 | P/B | 鱼产力(t) |
水生物维管束植物 | 3.03×104 | 25% | 100 | 75.75 |
3.8.2.4 水体有机碎屑渔业潜力评估
采用生物量转化法,根据有机碎屑有机碳年平均含量来计算碎屑类鱼类的年产量(表3-32),计算公式为:
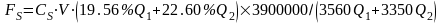
FS-碎屑食性鱼类或其它水生经济动物的增殖容量,单位为吨(t)
V-表层20 m以内的大水面容积,不足20 m的按实际容积计算,单位为108 立方米(108 m3)
CS-有机碎屑有机碳年平均含量,单位为毫克/升(mg/L)
Q1-水体中鲢占鲢、鳙的数量比例
Q2-水体中鳙占鲢、鳙的数量比例
种类 | 平均含量(mg/L) | 鲢鳙比 | V | 鱼产力(t) |
有机碎屑 | 15 | 2:1 | 1.704×108 | 586.85 |
3.8.2.5 小型鱼类渔业潜力评估
采用生物量转化法,根据西凉湖小型鱼类生物量计算出西凉湖食鱼性鱼类的鱼产力(表3-33),计算公式为:
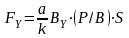
FY-鱼食性鱼类或其它水生经济动物的增殖容量,单位为吨(t);
BY-小型鱼类和虾类年平均生物量,单位为克/平方米(g/m2);
P/B-饵料生物年生产量与年平均生物量之比;不同区域湖泊和水库不同饵料生物的P/B系数
a-鱼类等水生经济动物对该类饵料生物允许的最大利用率;不同生态类型鱼类等水生经济动物对不同饵料生物允许的最大利用率
k-鱼类等水生经济动物对该类饵料生物的饵料系数,不同生态类型鱼类等水生经济动物对不同饵料生物的饵料系数
S-湖泊或水库面积,单位为平方千米(km2);
生物类群 | BY | P/B | S | 鱼产力(t) |
底层小型鱼类 | 7.5 | 2.5 | 85.20 | 79.88 |
上层小型鱼类 | 143.6 | 2.5 | 85.20 | 1529.34 |
3.8.2.6 西凉湖综合渔产潜力协同评估
根据西凉湖中水生生物资源现存量,计算出基于各类水生生物资源的综合渔产潜力,如表3-34。
类别 | 鱼产力(t) |
浮游植物 | 1663.61 |
浮游动物 | 2479.77 |
寡毛类 | 13.25 |
腹足类 | 25.26 |
水生昆虫类 | 178.06 |
水生物维管束植物 | 75.75 |
有机碎屑 | 586.85 |
底层小型鱼类 | 79.88 |
上层小型鱼类 |
西凉湖属于浅水型湖泊,东、西两湖区因中间水道搁浅而导致水交换率低,湖泊内源污染严重,藻类发生水华风险较高。2018、2019年西凉湖总氮、总磷含量较高,是导致藻类潜在发生水华的直接因素,并且西湖区较东湖区水生植物偏少,更容易引起潜在的藻类爆发。因此,为了降低藻类水华引起的潜在危险,保护西凉湖水生生物多样性,对西凉湖进行水生态修复与保护势在必行。
高等水生植物在水体中能够起到净化水体,为水生生物提供栖息地的生态功能。历史上,西凉湖是长江中游典型浅水草型湖泊,水生植物丰富,种类繁多,但随着围湖造田、围栏养殖、过度打捞等人为活动,水生植物的种类与数量急剧下降,目前,西凉湖水生植物较单一,大型沉水植物较少,水质净化能力较差,并且还影响其他水生物资源,如附着生存的大型软体动物,产粘性卵的鱼类等等。
水葫芦根系吸收能力强,能富集水中养分,在水中繁殖速度快,在湖面容易大面积覆盖。西凉湖水体富营养化严重,为水葫芦的生长提供了充足的养分,并成为当地的优势种,抑制或影响其他物种的生长,破坏生态多样性,极易造成湖区生态恶化,物种单一。因此,为了控制水葫芦在西凉湖区大面积入侵,防止造成生态系统不平衡的现象,对西凉湖进行水生态修复的工作必不可少。
鱼类是湖泊中的顶级消费者,通过营养级联效应作用于水体调控水质。目前,西凉湖的鱼类群落结构简单、资源量较小、种类单一,主要表现为:一是西凉湖滤食性鱼类如鲢、鳙较少,根据西凉湖水质状况,容易潜在爆发水华,而滤食性鱼类主要是摄食浮游植物和浮游动物;二是西凉湖鱼类多样性较低,主要以小型鱼类如似鱎、䱗为主,大型鱼类较少,且鱼类食性组成较单一,容易导致生态系统不平衡。适当的增加水体食鱼性如翘嘴鲌、鳜等能够有效的控制水体中小型鱼类的大量增殖,另外在水体中可以适当增加一些名品种,如黄颡鱼、中华鳖等,能够进一步优化鱼类群落结构。这与江湖阻隔、生境退化、围栏养鱼、过度捕捞和富营养化等过程紧密相关。
以习近平生态文明思想为指导,以建设生态文明环境为总目标全面贯彻落实《中共中央国务院关于加快推进生态文明建设的意见》(2015年4月25日,中发〔2015〕12号)、《国务院办公厅关于加强长江水生生物保护工作的意见》(2018年10月15日,国办发〔2018〕95号)、《国务院关于印发水污染防治行动计划的通知》(2015年4月2日,国发〔2015〕17号)、《农业农村部等10部委关于加快推进水产养殖业绿色发展的若干意见》(2019年2月15日,农渔发[2019]1号)、《农业农村部 生态环境部 国家林业和草原局关于推进大水面生态渔业发展的指导意见》(农渔发〔2019〕28号),持续推进西凉湖生态修复与保护,打造“绿水青山就是金山银山”创新实践基地。
坚持统筹规划、协调推进。紧密围绕国家生态文明建设目标,统筹空间布局、经济发展、环境保护、生态文化等多个维度,协调各有关行业、地方,发挥政府和市场作用,积极推动西凉湖生态文明发展,围绕生态文明建设标准的系统规划和协同开展。
坚持生态需求、突出重点。坚持节约优先、保护优先、自然恢复为主的方针;绿水青山就是金山银山,贯彻创新、协调、绿色、开放、共享的发展理念。建设西凉湖生态文明建设,深度进行环境治理。
坚持生态优先原则。生态文明建设是湖泊发展的基础,也是水生生物资源可持续发展的基础。良好生态环境是最普惠的民生福祉,坚持生态惠民、生态利民、生态为民。
坚持人与自然和谐共生。生态文明是人类赖以生存的基础,人类的生存离不开环境,同样生态文明是需要人们来共同建设,自然生产力是社会生产力的前提与基础,没有生态自然的宝贵财富,其他一切无法继续开展。
4.1.3.1 总体目标
通过对西凉湖进行水生态修复工程,持续改善湖泊水质,建立起适合西凉湖生态发展的体系,使西凉湖由富营养化湖泊逐渐转变成健康湖泊,让当地居民与湖泊发展形成一种互利共赢的模式,不断的优化和完善湖泊结构,提高水生物多样性,从而为西凉湖进一步发展提供了保障。
4.1.3.2 具体目标
(1) 减少周围居民生活污水、工业废水向西凉湖排放,降低湖泊富营养化程度,从而改善水质状况。
(2) 提高鱼类群落多样性,逐渐完善生态系统结构,提高生态系统的稳定性。
(3) 提高西凉湖渔业潜力,使湖区在发展的同时能够提高经济效益。
(4) 加强西凉湖水域治理,大力发展旅游业,带动湖区周围经济的发展。
本次规划涉及的水生态修复与保护的内容包含湖区周围环境治理、沉水植物的修复、挺水植物的修复、浮叶植物的修复、水葫芦的治理、湖泊软体动物的修复以及湖泊鱼类群落调控和藻类水华预防与处理八个部分。根据西凉湖生态环境、资源利用以及经济效益等方面,在西凉湖大规模的开展生态修复措施,解决西凉湖目前所存在的主要环境问题,根据西凉湖生态修复的目标,制定好明确的方案,确保生态环境能够得到最大可持续发展,合理开发渔业生产,维护国家生态,有序实施西凉湖生态修复项目。
西凉湖周围居民较多,生活污水、农业废水的排放加剧了水体富营养化程度,为保护西凉湖水域,改善水质状况,具体需要做到:
(一)周围居民在进行农业种植时,减少对化肥的使用,从而降低西凉湖水体氮、磷等营养元素含量。
(二)在湖区周围设置固定垃圾回收点,拒绝将生活垃圾直接进入湖区,造成湖区进一步污染。
(三)对污水处理厂的监管继续加强,重点监控污水排放是否达标,全面分析、掌控污水处理厂运行及污染物排放情况。
4.2.2.1 沉水植物修复目的
沉水植物是生态系统中重要的组成成分,在食物链和食物网中起初级生产者的作用。沉水植物可以吸收氮磷等物质用于生成自身的根系、茎叶等,同时沉水植物还可以用于黑臭水体的净化,对污染性水体的修复具有良好的促进作用。在藻类控制中,沉水植物可以通过根系吸收污染物,有相关实验证明狐尾藻、金鱼藻、眼子菜、黑藻等沉水植物可以通过化感作用抑制某些藻类的生长,防治水华的发生;通过根系微生物对污染物质进行降解、转化、分解,去除水体中的污染物质,同时沉水植物还能够通过光合作用释放氧气,为水中的生物生存提供了重要的基础保障。沉水植物群落的修复有助于增加生物多样性,有效的改善水体环境,最终达到净化水域的目的。
4.2.2.2 沉水植物种类选择
针对西凉湖水质环境特点,结合周围环境状况,对西凉湖大型沉水植物重建、种植需要考虑以下几点:
1.不同类型的沉水植物种子的萌发与激素、水环境条件、温度、光强以及氮磷相关,在进行沉水植物重建同时,考虑到该物种是否适合在西凉湖区中生长。
2.在水华暴发的富营养化水体中,由于藻类的快速生长一方面造成水体透明度下降,影响沉水植物的光合作用;另一方面造成水体无机碳、pH值剧烈变化,从而影响沉水的生长。
3.不同的水体底质对沉水植物种子的萌发也有显著影响,一般来说砂石底质不利于沉水植物的生长。
拟选取西凉湖水草分布比较薄弱的地方,并参考历史资料上曾经在西凉湖所生长过的沉水植物。主要种植的种类有轮叶黑藻、穗花狐尾藻、金鱼藻以及微齿眼子菜等植物,前期选择可以选择耐污性较强的穗花狐尾藻和轮叶黑藻在西凉湖水体扩大生态幅度;在具有了一定的生态范围后,可以选择种群扩张速度快,生物量较大的金鱼藻,可以迅速补充西凉湖内所缺乏的沉水植物;后期可以选择一些比较稳定的品种,如微齿眼子菜、马来眼子菜和苦草。水草的种植能够有效的降低水体中氮磷比,改善水体环境,降低富营养化程度,丰富生态系统的多样性。沉水植物还可以为水体内水生生物提供一定的栖息环境,有利于某些生物的生长和繁殖,沉水植物的恢复是水生态修复的重要保障之一。本次方案选择沉水植物主要有以下几种,具体特征见表4-1
金鱼藻
金鱼藻(Ceratophyllum demersum L.)是多年生沉水草本,多生于池塘、种植适宜水深<0.5-2.0 m,或 流速较缓的沟渠(图4-1)。金鱼藻耐污能力较强,对水体透明度需求较低,能够耐受富营养化水体的胁迫作用,通常是湖泊在富营养化进程中最后存留的沉水植物,其繁殖体很难收集,一般使用成年断枝或植株顶端部分进行种群重建。但是金鱼藻生物量年际变化较大,种群不稳定,并且由于根系不发 达,绝大部分生物量集中在植株顶端,无法形成对表层底质生境的改善; 此外金鱼藻植株极易折断,影响修复效果。因此金鱼藻一般用于较小的景观水体或极度富营养化水体生态修复中的先锋物种。


轮叶黑藻
轮叶黑藻(Hydrilla verticillata) 属单子叶多年生沉水植物(图4-2),本地湖泊常见的物种,茎长50-80 cm,叶带状披针形,4-8 片轮生,通常以4-6 片为多,长1.5 cm 左右,宽约1.5-2.0 cm。叶缘具小锯齿,叶无柄。轮叶黑藻冬季为休眠期,水温 10 ℃以上时,芽苞开始萌发生长,每年4 月开始进入营养生长阶段,5-8 月轮叶黑藻逐渐长达40-80 cm,达到最大生物量,能够在水体中形成巨大的“水下森林”现象。由于轮叶黑藻净化水体作用显著,对水生态系统结构和功能的稳定具有重要作用。因此,在富营养化水体植被恢复工程中被用作先锋物种。轮叶黑藻不仅具有良好的生态学价值,还具有营养学价值和综合价值。轮叶黑藻粗蛋白质含量高,占干重的 25.3%,而粗纤维和粗脂肪的含量均较低,是一种理想的高蛋白植物,其根、茎和叶都是草食性鱼类和河蟹的适口性青饲料。生长茂盛的轮叶黑藻可为河蟹、青虾等提供不可缺少的栖息和庇护场所,躲避敌害侵袭等。


穗花狐尾藻
穗花狐尾藻(Myriophyllum spicatum L.)为多年生沉水植物(图4-3),株高 1.0-2.0 m,种植适宜水深为0.3-2.0 m,以种子、越冬芽、根茎越冬,在南方部分地区也有以植株全体越冬。耐污能力较强,对水体透明度需求低,对水深耐受力较强,透明度深的耐受阈值接近 0.4 m。此外,穗花狐尾藻体内含有较高的酚类物质,适口性差,能够在草食性鱼类较多的水体中生存。同时,穗花狐尾藻根系较为发达,定植力强,并且穗花狐尾藻植株的抗拉力较强,能够承受较高的流速和风浪产生的剪切力,但是该物种生长速度较慢,种群扩张慢。因此,穗花狐尾藻可作为生境受损较为严重的湖泊中的水生植被修复种。


马来眼子菜
马来眼子菜(Potamogeton malaianus)是眼子菜科眼子菜属植物(图4-4),为多年生沉水植物,植株粗壮,移栽成活率较高,根系发达,具有很强的韧性,能够在流速较快的河流与风浪较大的湖泊中定植,适合在较深的水体中生长,但对水体透明度要求较高,种植适宜水深为0.3-1.5 m。马来眼子菜对水质有较好的净化作用,吸附氮磷效果非常好,同时也有一定景观功能,是沉水植物修复中的常用物种之一。


(5) 苦草
苦草属(Vallisneria)植物在中国分布有3个物种:苦草(Vallisneria natans)、刺苦草(Vallisneria spinulosa)、密刺苦草(Vallisneria denseserrulata),均为长江中下游流域的常见物种。株高 40-80 cm 不等,地下根茎横生,喜温暖,耐隐蔽,对土壤要求不严,种植适宜水深为 0.3-2.5 m,密集的苦草种群能够形成蔚为壮观的水下草甸,具有很好的景观效果(图4-5)。
苦草属植物对水下低光具有很好的耐受力,能够在透明度无法在短期 内有效改善的水体中开展修复,同时对水深耐受力较强,在某些湖泊中的分布水深可达6 m,并具有很强的水质净化能力,其通过地下匍匐茎形成分株的速度较快,种群扩张能力强。某些品种具有一定的耐寒能力,在长江中下游流域可保持叶片终年常绿,其中刺苦草容易形成大量的冬芽,具有很强的繁殖扩增能力。


种类 | 综合优点 | 综合缺点 | 生活类型 | 西凉湖最大种植水深 |
金鱼藻 | 1)污能力强 2)成活率高 3)种群扩张快 | 1)易形成高密度单优群落 2)生物量波动大 3)植株易断 | 多年生沉水 | 2.0 m |
轮叶黑藻 | 1)较耐低光 2)施工较方便 3)成活率较高 | 1)浅水区易形成高密度种群,影响景观 2)生物量波动较大 3)植株易断 | 多年生沉水 | 2.0 m |
穗花狐尾藻 | 1)耐污能力强 2)较耐低光 3)植株强壮 4)抗水生动物牧食 5)施工方便 | 1)景观较差 | 多年生或一年生 | 2.0 m |
马来眼子菜 | 1)植株强壮耐风浪 2)生物量较稳定 3)景观效果较好 4)管理方便 | 1)不耐低光 2)种植方式单一 3)成活率较低 4)耐污能力较弱 | 多年生沉水 | 1.5 m |
苦草 | 1)耐低光 2)生物量稳定 3)形态便于施工 4)稳定底泥效果好 5)景观效果好 | 1)易受水生动物牧食 | 多年生或一年生 | 2.5 m |
6)便于管理 |
4.2.2.3 修复区域与种植方式选择
依据现场调查以及考虑到湖区的水深以及丰水期和枯水期,在水深0-1.5 m以内、水体透明度较高、近岸地质适合、风浪较小的地区设置沉水植物恢复区。另外尽量选择天晴是进行沉水植物移植,刮风或起浪有可能会影响种植率。初期种植时用一定面积的围网将其保护起来,防止一些草食性的鱼类将苗种摄食,网目的大小根据水体中的鱼类体型而设置,一些活动性较强的鱼类可能由于扰动原因对水草有所影响,待沉水植物生长到一定数量或者能够在水中稳定生存就可以拆除围网。主要位于西湖区西南方向沿岸带两侧至水道两侧,东湖区由水道两侧至东南方向沿岸带两侧(图4-6)。
西凉湖底质类型大致主要分为硬底质、自然淤泥底质和两者结合类型的底质。沉水植物的根系无法在硬底质中定植,需先对底质利用生态基质材料改良,后采用无纺布包裹抛种法;自然淤泥底质中泥土较多,土质较松软,适合采用扦插法。

金鱼藻种植方法:金鱼藻的恢复措施大致有以下两种方法:1)每年5月份以后可在其它水域购买新长的金鱼藻全草进行移栽;此时移栽必须要用围网隔开,以防止水草随风漂走或被草食性鱼类破坏;围网面积一般在100-200 m2/个,每亩2-3个,每亩草用量100-200 kg,待水草落泥成活后可拆走围网。2)在湖区的一角或湖滨池塘设立水草培育区,专门培育金鱼藻。每年l0月份进行移栽,到次年4-5月份就可获得大量水草;培育区每亩用草量50-100 kg,每年可收获鲜草约5000 kg,可供25-50亩水面用草。培育区栽后管理主要有三个方面,一为水位凋节,金鱼藻一般栽在深水与浅水交汇处,水深不超过2 m,最好控制在1.5 m左右;二为水质调节,水清是水草生长的重要条件,建议先用生石灰调节将水调清,然后种草。
轮叶黑藻种植方法:1)播种时间在3月下旬至5月,水温上升至l5℃以上时便可播种,播种水底应在光补偿点范围内。播种前须用湖水浸种3-5天,洗去种粒的附着外皮,再用湿粘土拌种撒播。每亩用种量为150-250 g,播种后15天左右便可出芽。2)在4-5月,利用黑藻冬芽,在符合条件但没有沉水植物的水域撒播,亦能快速恢复黑藻种群。播种区域主要分布在沿岸区和水较深的区域。3)在每年的5-8月,轮叶黑藻已长成长达40-60 cm,可以在其它水域购买成熟植株,洒在淤泥较深但没有沉水植被的区域,每亩放草100-200 kg。该区域最好在枯水期选定,用围网与大湖区域分割开来,确保网隔内无草食性鱼类。
马来眼子菜种植方法:马来眼子菜最适宜的栽种季节为早春季节。常采用的栽种方法为扦插法:把马来眼子菜剪成10-15 cm一条,一株15条左右,全池按照横向3 m一株竖向2 m一株栽下。种完后上水至40 cm左右随后根据马来眼子菜生长情况加水。马来眼子菜生长过程中需时刻观察马来眼子菜生长情况,生长的过于宽大要及时捞取部分,防止高温期大面积死亡破坏水质,并影响景观效果。
穗花狐尾藻种植方法:穗花狐尾藻最适宜种植时间为每年的四至八月份左右。播种法:先挑选颗粒饱满且没有损害的种子,再将种子放置在干燥环境下保存一段时间,然后再将其种植下,种植水温要尽量保持在20-25℃。扦插法:剪取健壮的2-3节茎段,一般5-8厘米长,对茎段下部节上的轮生叶稍加修剪,插入基质中,深度约一半左右。插后管理主要是控制好温度与湿度,保持插床湿润和空气湿润,室温以20-25℃为宜,2周后便可移栽。
苦草种植方法:在4月中旬,水温回升至15℃ 以上,在沿岸带,选择水深小于30 cm、透明度较好的浅水区播种苦草种子,每亩播种量50 g,播种区域建议用网片围住。播种前选择晴朗天气,晒种1-2天,然后用湖水浸种3-5天,捞出后搓揉,搓出果荚内的种子,并将种子上的粘液清洗干净,再用半干半湿的细土或细砂拌种洒播。水温至18-22℃时,苦草种子经4-5天便开始发芽;至15天时出苗率便超过98%,30天后幼苗全长可达5-7 cm。
4.2.2.4 工程方案设计
根据西凉湖水体透明度和底质类型,选择穗花狐尾藻和苦草作为主要沉水植物修复种,同时搭配金鱼藻、轮叶黑藻和马来眼子菜,通过斑块镶嵌格局优化方式进行种植,苦草、轮叶黑藻、穗花狐尾藻、金鱼藻、马来眼子菜的种植面积比例为50:30:15:10:1,总种植面积为10.01 km2,工程量见表4-2。
种类 | 种植时间 | 工程量(km2) | 底质类型 | 种植方法 |
苦草 | 4月中旬 | 4.72 | 硬底质 | 抛种法 |
穗花狐尾藻 | 4-8月份 | 1.42 | 淤泥底质 | 扦插法、抛种法 |
轮叶黑藻 | 3月下旬-5月份 | 2.83 | 硬底质 | 抛种法 |
金鱼藻 | 5月份以后 | 0.94 | 淤泥或硬底质 | 营养体移植法 |
马来眼子菜 | 3月份 | 0.10 | 淤泥底质 | 扦插法 |
4.2.2.5 沉水植物修复工程技术分析
沉水植物群落修复工程中,常见的种植技术有扦插法、抛种法等,且每种种植方法均有不同的优缺点,对应的适用范围也有差异,常见的修复方式如表4-3。
修复方式 | 综合优点 | 综合缺点 | 适用性 |
单株扦插 | (1) 便于控制种群密度 (2) 便于植株固定 | (1) 效率较低 (2) 实施区域受水深限制 | 1.5 m浅水区 |
成簇扦插 | (1) 便于植株固定 (2) 成活率较高 (3) 便于控制种群密度 | (1) 准备时间长 (2) 实施区域受水深限制 | 1.5 m浅水区 |
单株抛植 | (1) 实施区域不受水深限制 (2) 适合种植较长的植株 (3) 施工效率高 | (1) 种群密度和范围不易控制 (2) 施工易受风浪影响 | 1-3 m区域 |
成簇抛植 | (1) 实施区域不受水深限制 (2) 适合种植较长的植株 (3) 施工效率高 (4) 成活率较高 | (1) 种群密度和范围不易控制 (2) 较易受风浪影响 | 1-3 m区域 |
扦插法的具体操作方法为:将预处理后的沉水植物种苗直接扦插入湖底土壤中,插入深度5-10 cm;当水深过大人工无法直接扦插时,可采用辅助工具进行扦插种植:将一节长1-1.5 m的竹竿顶端挖成 U 型缺口,取沉水植物种苗的根部嵌入至缺口中,再将竹竿缺口向下平稳快速插入湖底10 cm左右拔出。扦插种植主要在水深不超过 1.5m,淤泥或粘土底质的区域中实施。
无纺布包裹抛种法具体操作方法为:所选泥土,黏性较大的为佳。将植物的种子、芽孢完全裹入泥土,或将植物鲜体的一部分根茎裹入泥土并用无纺布包裹,缓慢抛入水中即可。
营养体移植法是主要以高等水生植物无性繁殖方式进行繁殖,利用营养体移植是成功率较高的方法,这些方法在湿地恢复工作中已经得到了广泛应用。
4.2.3.1 挺水植物修复目的
根据西凉湖挺水植物调查以及西凉湖水质综合分析的结果,湖区内挺水植物主要以菰(茭白)为主,物种多样性较低。西凉湖挺水植物群落修复有利于恢复生物多样性,同时还可以吸收并净化污水中的营养盐成分从而控制水体的富营养化,降低水体富营养化程度。同时挺水植物是连接水体和底泥沉积物物质循环的重要纽带,挺水植物扎根于底泥之中,以底泥为生长基质不仅能够实现水体与底泥中营养物质循环,降低沉积物中营养物质和重金属的含量,还能改变水体的动力学状态和降低水流冲刷作用。此外,挺水植物还能进行光合作用,产生的氧气通过自身通气组织输送至植物根系,继而向根系其他区泌氧,增加水体的溶氧量。高大的挺水植物还能为某些特性喜阴生物提供较好的栖息环境。因此,挺水植物群落的修复能够净化湖区水质,促进湖区的生态修复进程。
4.2.3.2 挺水植物种类选择
挺水植物能够增加生态系统的多样性,在参与物质循环以及能量转换同时还能净化水体,某些挺水植物如莲还具有食用价值。根据历史资料曾经在西凉湖出现过的挺水植物,选取耐污性强、成活率高、抗风浪的挺水植物,本次方案选择挺水植物主要有莲、水葱、芦苇、狭叶香蒲,挺水植物修复种类特点如表4-4。
(1)莲
莲(Nelumbo nucifera),又称荷、荷花、莲花、芙蕖、鞭蓉、水芙蓉、水芝、水芸、水旦、水华等,溪客、玉环是其雅称,未开的花蕾称菡萏,已开的花朵称鞭蕖,莲科,属多年生水生宿根草本植物,其地下茎称藕,能食用,叶入药,莲子为上乘补品,花可供观赏(图4-7)。是我国十大名花之一。根状茎横走,粗而肥厚,节间膨大,内有纵横通气孔道,节部缢缩。叶基生,挺出水面,盾形,直径30-90 cm,波状边缘,上面深绿色,下面浅绿色。叶柄有小刺,长1-2 m,挺出水面。花单生,直径10-25 cm,椭圆花瓣多数,白色或粉红色,花柄长1-2 m。花托在果期膨大,直径5-10 cm,海绵质。坚果椭圆形和卵圆形,长1.5-2.0 cm,灰褐色。种子卵圆形,长1.2-1.7 cm,种皮红棕色。生于池塘、浅湖泊及稻田中。中国南北各省有自生或栽培,经济价值高。人们习惯上称种子为"莲子"、地下茎为"藕"、花托为"莲蓬"、叶为"荷叶" 。


(2)水葱
水葱(Scirpus validus Vahl )属于莎草科藨草属,多年生挺水草本植物,高1-2 m,最佳生长温度15-30℃,10℃以下停止生长,能耐低温(图4-8)。生长在湖边或浅水塘中,适宜水深≤0.35 m。可采用播种、分株繁殖。在具有观赏价值的同时,对污水中有机物、磷酸盐及重金属有较高的除去率。


(3)芦苇
芦苇(Phragmites australis)是多年水生或湿生的高大禾草,生长在灌溉沟渠旁、河堤沼泽地等,世界各地均有生长,芦叶、芦花、芦茎、芦根、芦笋均可入药(图4-9)。芦茎、芦根还可以用于造纸行业,以及生物制剂。经过加工的芦茎还可以做成工艺品。古时古人用芦苇制扫把。芦苇是湿地环境中生长的主要植物之一。芦苇的根状茎十分发达。秆直立,高1-3 m,直径1-4 cm,具20多节,基部和上部的节间较短,最长节间位于下部第4-6节,长20-25 cm,节下被腊粉。芦苇具有横走的根状茎,在自然生境中以根状茎繁殖为主,根状茎纵横交错形成网状,甚至在水面上形成较厚的根状茎层,人、畜可以在上面行走。根状茎具有很强的生命力,能较长时间埋在地下,1 m甚至1 m以上的根状茎,一旦条件适宜,仍可发育成新枝。也能以种子繁殖,种子可随风传播。对水分的适应幅度很宽,从土壤湿润到长年积水,从水深几厘米至1 m以上,都能形成芦苇群落。在水深20-50 cm,流速缓慢的河、湖,可形成高大的禾草群落,素有“禾草森林”之称。


(4)狭叶香蒲
狭叶香蒲(Typha angustifolia) 是香蒲科香蒲属的植物,多年生,水生或沼生草本(图4-10)。根状茎乳黄色、灰黄色,先端白色。地上茎直立,粗壮,高约1.5-2.5 (-3) m。叶片长54-120 cm,宽0.4-0.9 cm,上部扁平,中部以下腹面微凹,背面向下逐渐隆起呈凸形,下部横切面呈半圆形,细胞间隙大,呈海绵状;叶鞘抱茎。雌雄花序相距2.5-6.9 cm;雄花序轴具褐色扁柔毛,单出,或分叉;叶状苞片1-3枚,花后脱落;雌花序长15-30 cm,基部具1枚叶状苞片,通常比叶片宽,花后脱落;雄花由3枚雄蕊合生,有时2枚或4枚组成,花药长约2 mm,长距圆形,花粉粉单体,近球形、卵形或三角形,纹饰网状,花丝短,细弱,下部合生成柄,长(1.5-) 2-3 mm,向下渐宽;雌花具小苞片;孕性雌花柱头窄条形或披针形,长约1.3-1.8 mm,花柱长1-1.5 mm,子房纺锤形,长约1 mm,具褐色斑点,子房柄纤细,长约5 mm;不孕雌花子房倒圆锥形,长1-1.2 mm,具褐色斑点,先端黄褐色,不育柱头短尖;白色丝状毛着生于子房柄基部,并向上延伸,与小苞片近等长,均短于柱头。小坚果长椭圆形,长约1.5 mm,具褐色斑点,纵裂。


种类 | 综合优点 | 综合缺点 | 生活类型 | 西凉湖最大种植水深 |
莲 | 1)污能力强 2)耐强光 3)观赏价值高 | 1)缺少淤泥影响存活 | 多年生挺水 | 1.0 m |
水葱 | 1)有一定耐寒能力 2)耐低温 3)景观效果好 | 1)阴雨天易受到病菌侵染 | 多年生挺水 | 0.35 m |
芦苇 | 1)净化水质 2)抗倒伏 3)短期成型 4)生物量较高 5)适应性强 | 1)对渠道供水输水有影响 2)减缓水的流速 | 多年生挺水 | 1.0 m |
狭叶香蒲 | 1)喜热耐旱 2)适应性强 3)喜肥耐贫 | 1)再生能力差 2)易形成单优势种群落 3)适口性较差 | 多年生沼生 | 1.0 m |
4.2.3.3 修复区域与种植方式选择
根据目前西凉湖水体透明度、沿岸带类型以及水质条件等多方面综合分析,选择湖岸带有弯折并且风浪较小的区域进行种植,主要位于西湖区西南方向沿岸带两侧至水道,东湖区由水道两侧至东南方向沿岸带两侧(图4-11)。
莲种植方法:莲喜相对稳定的静水,忌涨落悬殊和风浪较大的流水,水深一般不宜超过1.5 m。生长季茎叶最适温度为25-30 ℃。要求日照充足,不宜长期在室内栽培。土质以富含有机质的粘壤土为宜。莲子寿命特别长,千年古莲子仍能萌发新株。莲以分株繁殖较常用,也可播种繁殖。均宜在春季转暖后进行。栽种时,可采取藕节繁殖方法和顶芽繁殖方法进行扦插,株行距一般保持在200×200 cm到400×200 cm。
狭叶香蒲种植方法:分株法:一般在气温达到15 ℃以上时,将狭叶香蒲地下的根状茎挖出、洗净,用利刀截成带3-5个芽的茎段,分别定植。宜在4月中旬选取苗种种植,种植时选取每丛带3-5个芽且生长健壮的种苗插入泥中,栽植深度一般10-15 cm,株间距保持约50 cm。播种法:多于春季进行,播种前先进行催芽处理,然后播于苗床,注意保持苗床湿润,夏季小苗成形后移栽。
水葱种植方法:早春天气渐暖时,把越冬苗从地下挖起,抖掉部分泥土,用枝剪或铁锨将地下茎分成若干丛,每丛带5-8个茎杆;栽到无泄水孔的花盆内,并保持盆土一定的湿度或浅水,10-20天即可发芽。如作露地栽培,每丛保持8-12个芽为宜。露地栽培时,于水景区选择合适位置,挖穴丛植,株行距25 cm×36 cm,如肥料充足当年即可旺盛生长,连接成片。定植与株行距4-5月可移苗定植,为便于管理,最好分级栽植。株行距20 cm×20 cm,每穴3-4株,分蘖力强的品种株行距20 cm×25 cm,每穴2-3株。定植时用尖圆柱形木棒插孔,栽植深度以露心为宜。
芦苇种植方法:芦苇具有横走的根状茎,在自然生境中以根状茎繁殖为主,根状茎纵横交错形成网状,甚至在水面上形成较厚的根状茎层,人、畜可以在上面行走。根状茎具有很强的生命力,能较长时间埋在地下,1 m甚至1 m以上的根状茎,一旦条件适宜,仍可发育成新枝。也能以种子繁殖,种子可随风传播。对水分的适应幅度很宽,从土壤湿润到长年积水,从水深几厘米至1米以上,都能形成芦苇群落。在水深20-50 cm,流速缓慢的河、湖,可形成高大的禾草群落。采取分根移栽法:在每年3月下旬至4月上旬,用铁锹在靠近苇苗处挖出长宽各15 cm,高20 cm左右的土佗,每个土佗上约有2-4株苇苗,按株行距均为1 m移栽。
4.2.3.4 工程方案设计
据西凉湖水体透明度和底质类型,选择莲、狭叶香蒲、水葱为主要挺水植物修复种,同时搭配芦苇,种植比例为6:14:1:9,总种植面积为10.01 km2,工程量见表4-5。
种类 | 种植时间 | 面积(km2) | 底质类型 | 种植方法 |
莲 | 3-5月份 | 2.00 | 淤泥底质 | 扦插法 |
狭叶香蒲 | 4月份中旬 | 4.67 | 淤泥底质 | 播种法、分株法 |
水葱 | 3-4月份 | 0.33 | 淤泥底质 | 分株法 |
芦苇 | 3月下旬-4月上旬 | 3.00 | 淤泥底质 | 分根移栽法 |
4.2.3.5 挺水植物工程方案分析
土壤:应该选择通气性好,且含有大量的腐殖质、疏松、肥沃的土壤,pH 值以 6.0-8.5 之间为宜、种植土厚度不小于 30 cm。
水深:挺水植物因茎叶会挺出水面,须保持50 cm至100 cm左右的水深。
种植方式:挺水植物的种植方式以扦插种植为主。首先在挺水植种植期间适当控制水位或选择水位较低的时间。其次在种植区内开挖深度10-20 cm的种植沟,种植沟间距1 m。最后,使用黏土对种植沟进行回填,恢复其原有高程。后期进行管护,及时扶正倒伏种苗,补种死亡种苗。
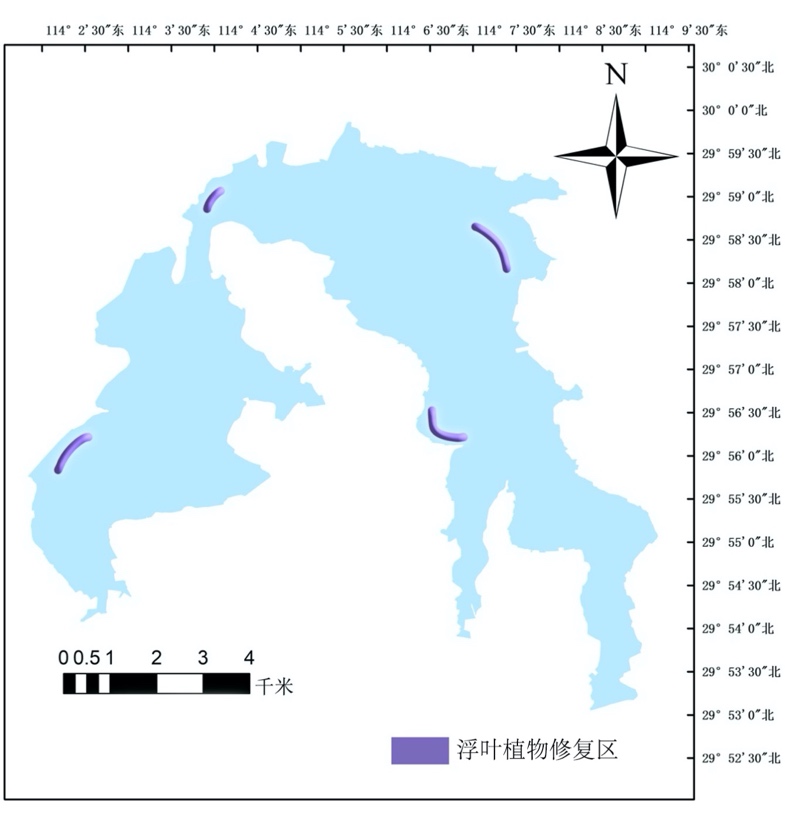
4.2.4.1 浮叶植物修复目的
浮叶植物生于浅水中,根长在水底土中的植物,仅在外表面有气孔,叶的蒸腾非常大。浮叶植物能够通过根部的吸收、吸附作用,削减富营养化水体中氮磷及有机质,能净化水体,还能够为水体中的鱼类提供饵料。浮叶植物能进行光合作用,为水体增加氧气,为其他水生生物提供必要的氧气保障。根据西凉湖浮叶植物调查以及西凉湖水质综合分析的结果,浮叶植物主要以菱角为优势种,且菱角分布区域广泛,生物量较大,群落多样性较低。通过修复浮叶植物群落结构,增加复叶植物多样性,有助于水体营养物质和重金属元素的去处,达到最终净化水域的目的。
4.2.4.2 浮叶植物种类选择
根据西凉湖水质状况以及修复区域底质类型,选择适合在西凉湖生长、耐污性较强、生物量较稳定的修复物种,主要有芡实、水鳖、荇菜,浮叶植物修复种类特点如表4-6
(1)芡实
芡实(Euryale ferox),又名鸡头米,为浮叶植物,睡莲科芡属一年生水生植物,原产东南亚,我国主要分布在长江中下游地区(图4-12)。在中国式园林中,与荷花、睡莲、香蒲等配植水景,尤多野趣。芡实种仁可供食用、酿酒。根、茎、叶、果均可人药。外壳可作染料。嫩叶柄和花柄剥去外皮可当菜吃。全草可作绿肥,煮熟后又可作饲料。芡实补中益气,为滋养强壮性食物,其味甘涩平,具有固肾涩精、补脾止泄的功效。芡实种仁含有丰富的蛋白质、碳水化合物、维生素及微量元素等,可作优良滋补品;种子外壳可作染料;叶柄和花梗去皮后鲜嫩清香,可作蔬菜食用;根茎富含淀粉,可作饲料。芡实是一种深受群众喜爱的绿色食品,营养价值高,种植成本小,利润大,生产管理简单。芡实一般每667 m2产鲜芡米50-60 kg,干芡米25-30 kg,叶柄、花柄每667 m2产量可达1500 kg,收入达3000 元左右,经济效益可观。种植芡实,为农业结构调整、农民增收提供了新的途径,市场前景非常好。


(2)水鳖
水鳖(Hydrocharis dubia),水鳖科水鳖属植物,多年生漂浮植物,我国广泛分布,通常在湖浅 水区漂浮生长,适宜水深 0.5-1.0 m(图4-13)。叶片圆状心形,背面有发达的气囊, 花白色,具有一定的景观效果。水鳖不受水体透明度限制,对富营养化水 体耐受较好,生境耐受力强,适合在小型池塘、景观水体中使用。可通过整株投放的方式进行栽培,需要对种群规模进行管理,防止泛滥。


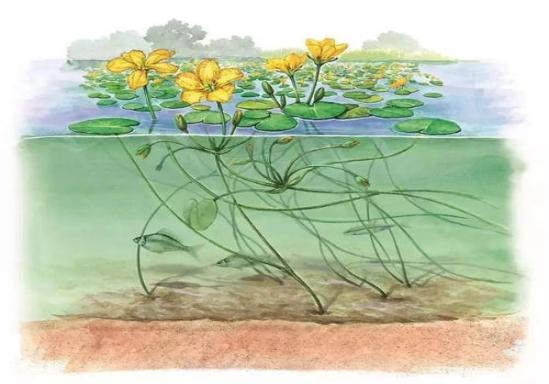
(3)荇菜
荇菜(Nymphoides peltatum),龙胆科荇菜属,为多年生浮叶植物(图4-14),在我国南北方均有广泛分布,适宜水深≤1.5 m,叶型优美,花黄色,具有很好的景观效果,对水体透明度要求不高,能够耐受一定程度的富营养化,种群密度适中,管理方便,适合作为伴生种提升植被景观效果。一般通过整体移栽的方式进行种植。

种类 | 综合优点 | 综合缺点 | 生活类型 | 西凉湖最大种植水深 |
芡实 | 1)有一定净水能力 2)具有一定观赏价值 | 1)不耐寒、不耐旱 | 一年生大型水生草本 | 1.0 m |
水鳖 | 1)净水能力较强 2)景观性高 | 1)休眠期较长,发生期也较长 | 多年或一年生 | 1.0 m |
荇菜 | 1)适应力强 2)景观性高 3)耐寒、耐热 4)净化水质 | 1)易形成单优势种群落 | 多年生浅水性植物 | 1.5 m |
4.2.4.3 修复区域与种植方式选择
根据目前西凉湖水体透明度、沿岸带类型以及水质条件等多方面综合分析,浮叶植物一般选择肥沃、疏松的土壤,pH以6.0-8.5为宜,种植土厚度不小于30 cm,并且对水质要求pH以6.0-8.5为宜,适宜在水面相对静止的围和合区域种植。由于浮叶植物茎较小,不仅容易受风浪影响而且水体中鱼类活动也对植物幼苗有影响,在种植过程成,可以通过围栏的形式,按照水体中鱼类体型的大小,设置围栏的网眼大小,待浮叶植物能够在水体稳定后可适当拆除围栏,修复区域主要位于西湖区西南方向沿岸带两侧至水道,东湖区由水道两侧至东南方向沿岸带两侧(图4-15)。
芡实种植方法:芡实种植的适宜土质以壤土最好,黏土次之,砂土最差。宜选择水位容易控制、肥土层较厚、有机质丰富的湖区。选择30-80 cm水深的场地对芡实进行栽种,在适宜芡实生长的前提下,方便收获芡米以及以收获鲜茎秆。4月上旬将上一年保存的种子捞起,清洗干净后进行粒选。选色深、无损伤的种子进行催芽。催芽时每盆放置种子的厚度不宜超过30 cm,加水淹没种子。盆口加盖塑料薄膜增温,有条件的晚上将催芽盆搬到室内保温,保证温度白天在20℃以上,晚上不低于5℃。到第4天至第5天换一次清水,50%左右的种子露白时即可播种。大面积湖荡栽培多直播。直播方法是将种子用泥团包裹,每团2-3粒,种植密度一般按株行距 100-200 cm进行。
水鳖种植方法:水鳖喜欢温暖湿润的环境,温度的高低会对水鳖草的生长有直接的影响。水鳖的种植应选择水位适宜, 30-80 cm的水位,并且在阳光充足的区域。养殖水鳖草时应该先在花盆中放好塘泥,加入充足的水分,再把水鳖草的茎剪断,直接插入到塘泥中,再放上水,等十几天左右就能看到新株长出,这时再把它们移动到小池中养殖,池中一定要施足底肥,而且不能让植株过密,不然会发生黄叶或者烂根等不良症状。
荇菜种植方法:荇菜一般生长在比较浅的水中,是我国分布十分广泛的一种野菜。荇菜的繁殖方法分为油性繁殖和无性繁殖。有性繁殖指的是种子繁殖,而无性繁殖则是指分株分株和扦插繁殖。种子繁殖一般在三月份进行催芽,四月初温度回升到十五度左右的时候进行播种。播种的时候要将种子均匀的撒在田间,然后将种子轻轻的拍入土壤中即可。分株繁殖一般是在每年的三月份进行,将生长过密的植株进行分割,然后形成单独的植株进行栽种。扦插繁殖只要选在气候条件较好的时间进行,选用荇菜的茎叶,要保留三四节,然后扦插入田中即可。
4.2.4.4 工程方案设计
根据西凉湖水体透明度和底质类型,选择荇菜、水鳖、芡实为主要修复种,种植比例为15:5:1,总种植面积为6.00 km2,工程量见表4-7。
种类 | 种植时间 | 面积(km2) | 底质类型 | 种植方法 |
荇菜 | 4月份 | 4.29 | 淤泥底质 | 抛种法 |
水鳖 | 4月份 | 1.43 | 淤泥底质 | 抛种法 |
芡实 | 4月上旬 | 0.29 | 淤泥底质 | 栽种法、抛种法 |
4.2.4.5 浮叶植物工程方案分析
土壤要求:选择肥沃、疏松土壤,PH 以6.0-8.5为宜。种植土厚度不小于30 cm
水质要求:水质 pH 以6.0-8.5为宜。盐碱地区种植时含盐量应控制在1.5‰,成活后含盐量可放宽至 2.0‰,NH3-N含量高的水体不适宜种植小型漂浮植物,可选择一些大型漂浮植物种植。
种植方式:浮叶植物种植采用带土抛植的方式进行种植。将符合长度标准的种苗根部使用粘土进行包裹,外层使用可降解的无纺布种植袋进行包裹,每3-5株组成1簇,装入1个种植袋中。实施中将制作好的抛植种苗缓慢放入水中,保证叶片在水面完全展开。
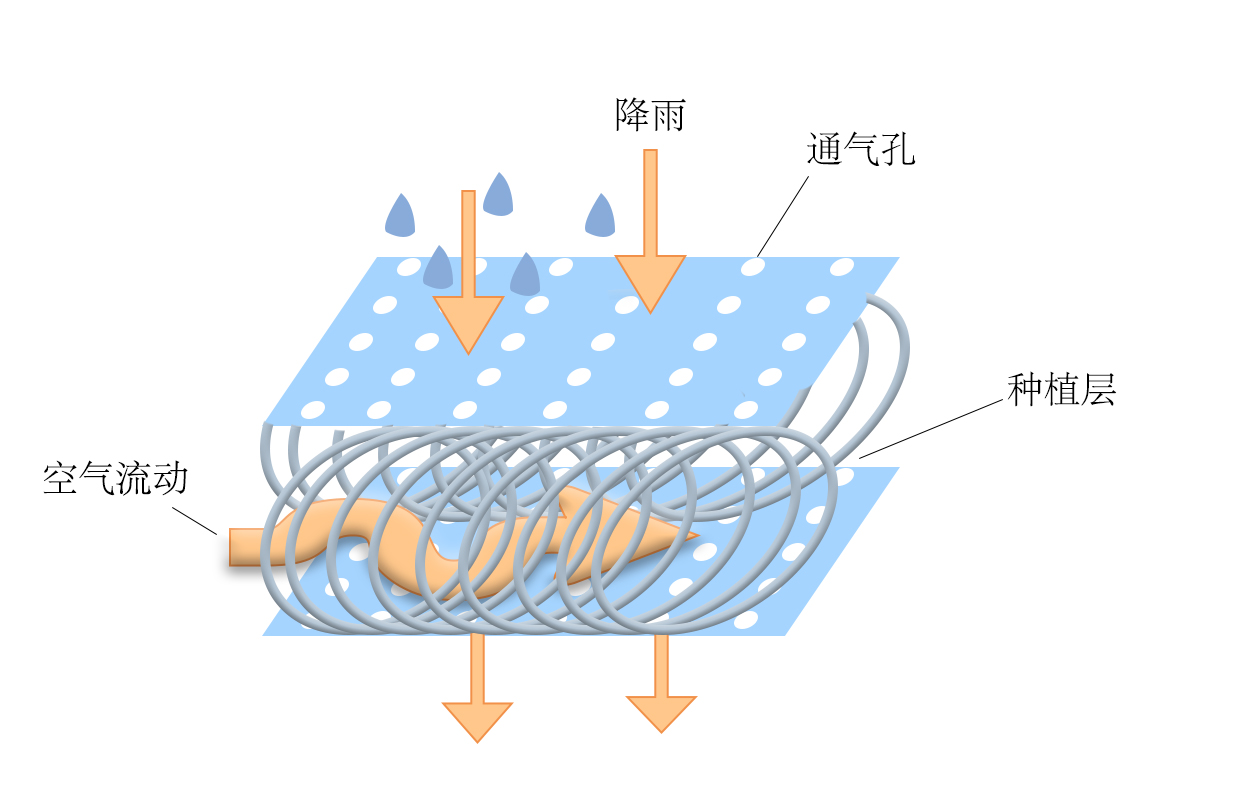
立体生态平台是一种净水系统,在去除污染物的同时并且能够达到净水,维持水环境健康的设施。在西凉湖污染较为严重的区域设置该平台,并在平台上种植具有净水且还有一定经济价值的水生高等植物(挺水植物如美人蕉等),植物的根部具有吸附作用,能够吸收水体过多的营养成分,再与平台上微生物共同作用,达到改善水质的作用。浮台上具有一定数量的孔,能为微生物的生长繁殖和截留污染物提供巨大的附着面积(图4-16)。
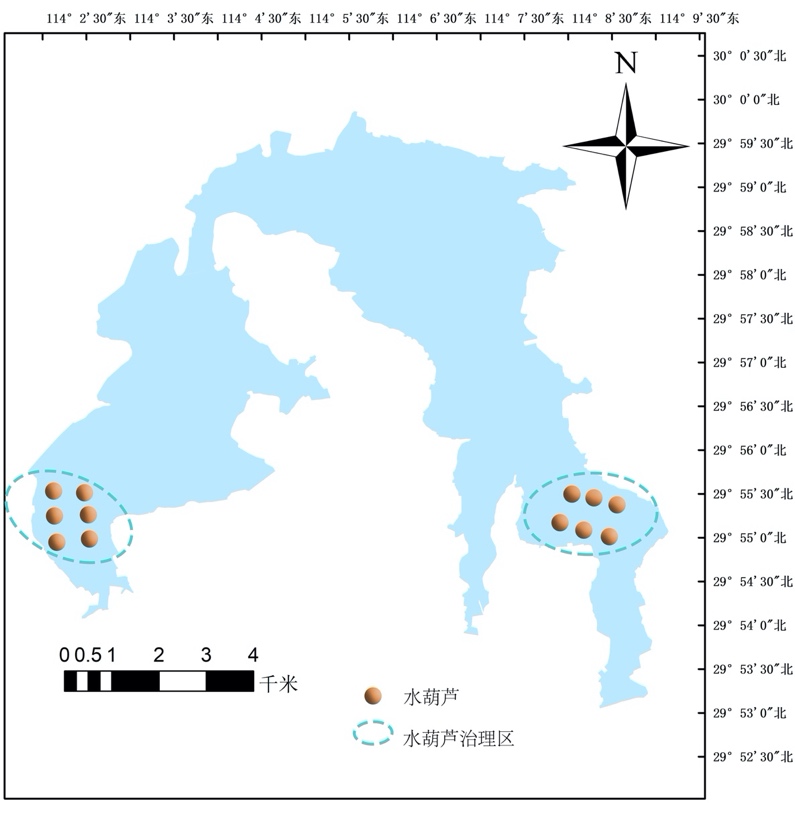
水葫芦,学名凤眼莲,属于雨久花科凤眼莲属植物,水葫芦茎叶悬垂于水上,蘖枝匍匐于水面。花为多棱喇叭状,花色艳丽美观。水葫芦十分喜肥,尤其是氮肥,水层养分含量高时,植株高大,根系短,开花少,繁殖快。水葫芦存活的温度范围也很广泛,在0-40℃都能够存活,其中在25-32℃时生长最快。水葫芦入侵性强,繁殖迅速,生长力旺盛,水葫芦有着较宽为生态幅,同时具有强大繁殖能力,能够利用各种营养资源,迅速形成大量的种群,大面积覆盖在湖泊上。
水葫芦的危害主要是大量繁殖所导致的,主要包括破坏水生生物多样性,影响水环境质量、阻碍行洪、影响航道、影响水体景观等问题。西凉湖属于富营养型湖泊,其水体性质很适合水葫芦的生长,因此,西凉湖水葫芦泛滥是当前湖泊迫切需要解决的问题。
4.2.6.1 水葫芦治理目标
水葫芦的治理目标主要是通过物理防治、化学防治以及生物防治等,对水葫芦生物量和密度进行控制,从而使水体恢复健康,使水生物资源减少受水葫芦的影响。
4.2.6.2 水葫芦治理方案
水葫芦治理主要有物理打捞、化学药物防控以及通过生物学效应来达到抑制水葫芦的生长(图4-17)。本次规划以生物学效应来达到治理水葫芦泛滥,生物学治理主要是通过草食性鱼类的摄食作用(如草鱼),草鱼在抑制水葫芦的同时还能够将其作为自身的饵料生物,来生长发育。具体实施方案有:
(一)人工围栏区设置:如图4-5所示在西湖区和东湖区呈“凹”字地形区域进行人工围栏,围栏面积为800 hm2(1.2万亩),将湖区中的水葫芦集中到“凹”字区域内(表4-8)。
(二)草鱼规格:选取健康无病害、品质规格较好的草鱼,规格为300-500 g(表4-9)。
(三)草鱼的投放:
根据调查,水葫芦密度为1893.76 g/m2,并由水葫芦现存量计算出鱼产力(表4-10),鱼产力计算公式参考3.8.2.3。
围栏时间 | 围栏面积(km2) | 围栏方式 | 网眼大小 |
6-9月份繁殖期后 | 8 | 固定地点集中围栏 | 不大于当年放养草鱼规格 |
种类 | 规格 | 密度 | 放养时间 |
草鱼 | 300-500 g | 900尾/km2 | 集中围栏处理后 |
种类 | 治理面积(km2) | 密度(g/m2) | 现存量(t) | 鱼产力(t) | |
水葫芦 | 8 | 1893.76 | 5.6×103 | 14.2 | |
根据西凉湖草鱼生长特性,估算草鱼年增重为原体重4倍,结合鱼产力、放养规格,计算出水葫芦治理区放养草鱼数量约为7200尾。
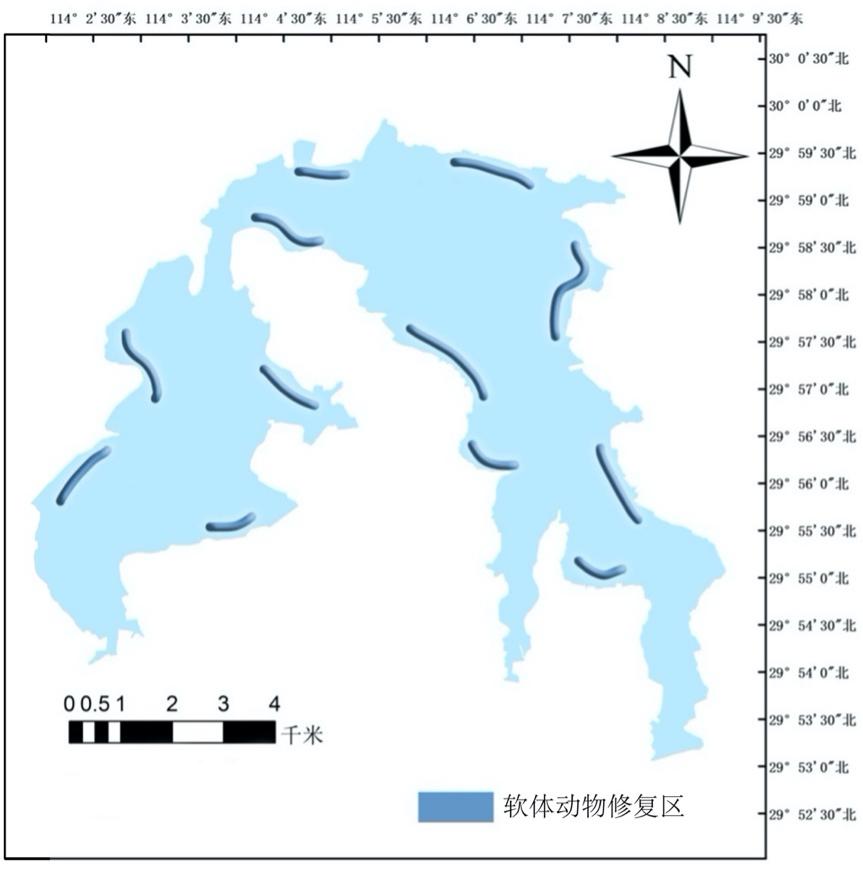
4.2.7.1 软体动物修复目的
大型软体动物在水生态系统物质循环与能量流动中有着特殊的地位和作用,可以滤食或摄食底栖藻类有效调节底泥-水界面营养盐的释放,软体动物在湖泊中以蚌和螺为主,这类生物能够絮凝水体悬浮物,滤食水体浮游植物,刮食附着藻类,并几乎不牧食水生植物。螺类如铜锈环棱螺与苦草共同作用能够降低水中总氨氮浓度、附着藻类和底栖生物量,二者结合在一起能够取得更显著的水体修复效果,通过调整浮游生物结构和种群的结构使水体环境朝有利的方向发展。
4.2.7.2 软体动物种类选择
根据西凉湖水质以及底泥特征,选取耐污性较强的修复物种,其中螺类主要使用环棱螺,搭配大沼螺、纹沼螺和萝卜螺投放;蚌类主要选择褶纹冠蚌,配合河蚬、无齿蚌、圆顶珠蚌投放。
(1)环棱螺属
环棱螺属(Bellamya)是腹足纲的一属,螺壳圆锥形,螺环面近于平;体环大,具旋棱;壳口卵圆形,口缘薄,上端角状,脐小。以水生植物嫩茎叶、细菌和有机碎屑为食。较常见的主要铜锈环棱螺和方形环棱螺(图4-18)。


(2)褶纹冠蚌
褶纹冠蚌(Cristaria plicata)隶属软体动物门,瓣鳃纲,蚌科。属大个体淡水贝类,壳大, 长近 30 cm,宽 10 cm,高 17 cm,呈不等边三角形,前背缘突出不明显,后背 缘伸展成巨大的冠。耐污水和低氧能力较强,喜栖于较肥的水域,生活在 硬底或泥沙底的河流、湖泊、沟渠等水域中,它比三角帆蚌分布广泛,在 我国几乎各地都出产(图4-19)。


4.2.7.3 软体动物修复区域
根据西凉湖水质状况与底栖生物分布情况综合分析,选择底质较好的区域进行投放,软体动物的恢复区域主要位于西湖区西南方向沿岸带两侧至水道,东湖区由水道两侧至东南方向沿岸带两侧(图4-20)。
4.2.7.4 工程方案设计
结合西凉湖软体动物分布情况以及历史资料,拟定西凉湖双壳类如褶纹冠蚌、河蚬,投放密度为 50 g/m2-60 g/m2,腹足类如环棱螺投放密度为 30 g/m2-50 g/m2。螺类和蚌类生活在水体的底层,其存活状况不易观察,定期的进行管理和维护,防止已经死亡的螺类和蚌类对水体造成污染。
螺类和蚌类投放时间考虑在繁殖期夏季或冬季水位较低时进行投放。投放时间与密度如表4-11所示
物种类型 | 投放时间 | 投放地点 | 投放面积(km2) | 投放密度 | ||
螺类 | 繁殖期夏季或冬季水位较低时候 | 近岸水深 1.5m 以内区域 | 6.6 | 30 g/m2-50 g/m2 | ||
蚌类 | 繁殖期夏季或冬季水位较低时候 | 近岸水深 1.5m 以内区域 | 6.6 | 50 g/m2-60 g/m2 | ||
根据历史资料,西凉湖鱼类资源丰富,种类繁多,但由于环境的改变,导致生态环境变差,鱼类所生存的场所被破坏,鱼类无法进行最大化的繁殖,导致鱼类多样性降低,甚至一些珍贵的鱼种也逐渐消失。湖泊栖息地的修复主要是通过人工鱼巢的方式为鱼类提供更多的繁殖场所,提高鱼类的产率,进而提高鱼类群落多样性。
在水体中设置复合型人工鱼巢,可以在鱼类繁殖季节,为草黏性卵、沉粘性鱼类提供产卵场所,对增养天然湖泊水生生物资源,维持水生生物多样性,保护水域环境,达到生态效益双丰收。具体设置鱼巢的方式是沿着湖岸边进行布置鱼巢,尽量选择比较空旷的地方。
人工鱼巢制作:
人工鱼巢的材料主要是由棕榈片构成,在制作时,先用小锤轻轻将棕榈片捶打片刻,然后多扯动几次,让它充分松软,目的事增加卵的附着面积。然后用铁圈将棕榈片固定住,4-5张棕榈片捆成一个铁圈,最后用浮球和绳子将每一个含有棕榈片的铁圈串联起来(图4-21、图4-22)。
技术分析:
通过在水体中设置人工鱼巢,能够味水体中多种粘性卵鱼类提供栖息庇护以及繁殖的场所,可用于西凉湖中鱼类以及其他水生生物资源的增殖,维持生物多样性,保护水域环境,提高经济效益。
人工鱼巢位置:
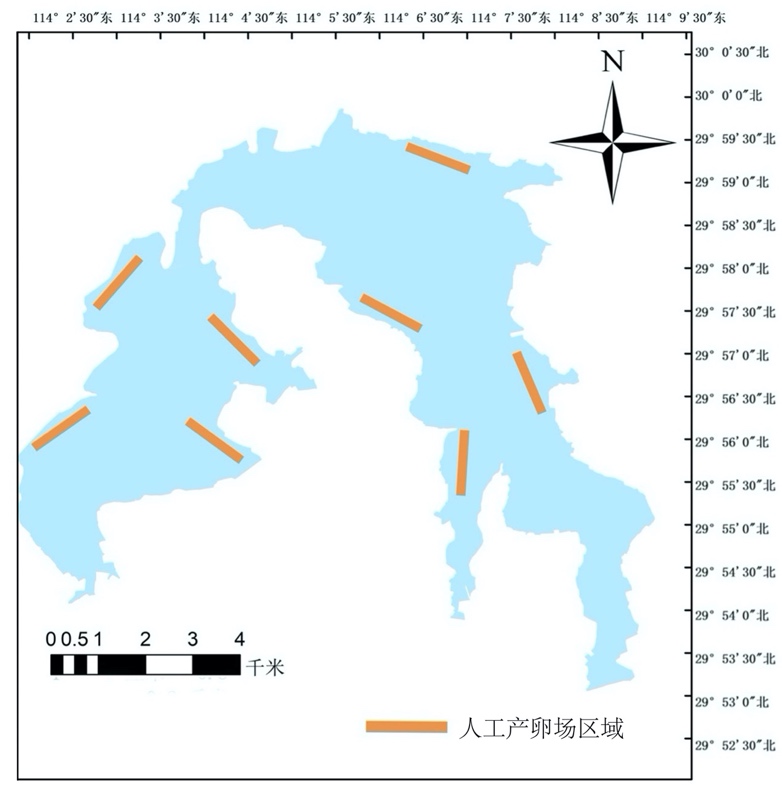
人工鱼巢安放的位置一般在湖泊沿岸,水位不宜太浅,尽量选择比较空旷、周围环境良好且不具有大风大浪的区域(图4-23)。本项目共建立8个区域,每个区域为1 hm2。
人工鱼巢设置时间:
一般人工鱼巢放置的时间在鱼类繁殖期之前,每年4月份是鱼类开始产卵的时间,所以鱼巢一般也在4月初左右进行安放。


鱼类在生态系统中起着重要的调控作用,是生态系统中的消费者,根据食物网、食物链的作用,既能够通过上行效应控制浮游动物的密度,又能够通过下行效应控制浮游藻类的密度。鱼类在生态系统中的摄食、活动都会对水体造成一定的影响,不同食性的鱼类在水体中摄食的食物种类不同,不同生活习性的鱼类在水体中活动的范围也不同。
调整鱼类群落结构有利于西凉湖水生态修复的进行,根据鱼类与浮游藻类之间的关系,有利于控制浮游藻类的密度与生物量,能够有效的减少水体出现水华的状况。
根据西凉湖鱼类群落调查,鱼类群落多样性较低,鱼类种群逐渐单一化,小型化。主要是有䱗、似鱎构成,而这类鱼在水体中繁殖快,并且数量多,很容易形成优势种,另外这两种鱼都是以浮游生物为食,在一定程度上,与鲢、鳙形成一定的竞争关系,不仅鲢、鳙对食物的利用,还减缓了鲢、鳙鱼的生长。我们要合理的调整手段,优化和调整鱼类群落结构,实现西凉湖水生态修复与保护。
4.2.9.1 鱼类增殖放流方案
根据不同水生生物资源的渔产潜力计算结果,制定鱼类增殖放流方案。西凉湖2021-2025年各种类放流时间、规格以及数量见表4-12,根据鱼苗的来源、市场、捕捞情况以及湖泊的状况可作出实际调整。
(1)滤食性鱼类增殖放流
湖泊中浮游植物的繁衍生长既可消耗水体中大量氮、磷元素,又为浮游动物提供饵料基础。如果水体的营养水平很高,体积大的蓝藻水华发生严重,这时可利用鲢、鳙这样的滤食性鱼类直接牧食蓝藻,消除蓝藻水华的堆积与危害。鲢、鳙是典型的滤食性鱼类,主要以水中的浮游植物、浮游动物及颗粒有机碎屑为食。譬如,在武汉东湖,利用鲢、鳙成功地控制蓝藻水华的发生超过30余年,有效控制蓝藻水华发生的鲢、鳙养殖密度约为50 g/m3。滤食性鱼类能够有效的改善生态环境,降低水体中浮游生物的密度,有效的抑制了水华的发生。根据水生态修复与保护原则,加上基于西凉湖浮游生物和有机碎屑的供饵能力及同类湖泊的经验,确定鲢、鳙的生态放养方案如下:
放养时间:鲢、鳙鱼种放养时间宜在每年大捕捞结束后的1-2月,宜在每年的6-7月补充放养夏花。投放鱼种应选择晴好天气进行,放养地点宜选在敞水区,远离进出水口。
放养规格和密度:鲢、鳙鱼种放养密度可根据鳙的放养规格、湖泊营养类型和预期上市规格确定,可根据放养后鲢、鳙生长和存活情况及上市规格要求作适当调整。为防止水华事件的发生,同时达到水生生物资源养护和生态修复的目的,对滤食性鱼类放养规格和密度作出以下建议:鲢、鳙的鱼苗放养可以分为三种规格:大规格鱼种,0.50 kg/尾;中规格鱼种,0.20 kg/尾;寸片鱼种,约100尾/kg。其中寸片鱼种或夏花鱼种可在夏季根据湖泊中鱼苗存活和生长情况进行补充。冬季放养规格可根据市场供应情况适当调整,规格0.50 kg/尾时,可增加投放量10%。应调整鲢、鳙的放养比例,加大鲢的放养、减少鳙的放养,保持鲢、鳙放养比例为2:1。从2021年开始鲢、鳙的放养量会按照浮游生物饵料资源状况进行调整。结合西凉湖浮游生物资源现状以及生物环境状况,建议2021-2025年鲢、鳙的放养规格、数量如下表4-13所示。根据每年鱼种资源和湖泊本底情况,2022-2025年放养的数量可做实际调整。
鲢(Hypophthalmichthys molitrix)
生括习性:性活泼,喜跳跃,生活于水的中上层。主要以浮游植物为食。最小成熟个体为3龄鱼,生殖时期为4月下旬至6月。产卵要求在有急流泡漩水的河段进行。卵沉性,受精卵卵膜吸水后膨胀,随水漂流发育。分布在长江、汉水干流以及湖泊(图4-24)。

鳙(Aristichthys nobilis)
生活习性:喜生活于水的中上层,动作较迟缓,不喜跳跃,易捕获。主要以浮游动物为食,亦食部分浮游植物。性成熟年龄为 4-5 龄,生殖时期在5月上旬至7月。要求在河流上游急流、有泡漩水的江段产卵。卵沉性,受精卵卵膜吸水膨胀。随水漂流发育。分布在长江、汉水干流,湖泊、水库(图4-25)。

(2)碎屑食性鱼类增殖放流
黄尾鲴和细鳞鲴是碎屑食性经济鱼类,主要摄食着生藻类和植物碎屑,具有生长迅速、抗病性强、捕捞方便等优势。由于食性和生活习性等特点,黄尾鲴和细鳞鲴在水体中有与其他鱼类不同的摄食来源,而根据西凉湖具有丰富的有机颗粒等,适合开展放养黄尾鲴和细鳞鲴,并且同时还能够改善水体的生态环境。根据湖泊特点以及结合当时市场的需求,基于水体供饵能力及同类湖泊的经验,确定的生态放养方案如下:
放养时间:夏花在当年6月份放养,冬片在冬季捕捞结束或次年3月份前放养。放养地点应远离进出水口,宜选择饵料资源丰富的水域。
放养规格和密度:夏花规格3-4 cm;冬片规格10-25 g。夏花鱼种放养密度5-15尾/亩,或冬片鱼种放养密度1-6尾/亩。结合西凉湖碎屑食性鱼类饵料生物资源现状,建议2021-2025年碎屑食性鱼类的放养规格、数量如下表4-13所示。
黄尾鲴(Xenocypris davidi)
生活习性:常生活于水的中下层,以下颌发达的角质边缘刮取岩石和泥底的固着藻类为食,亦食部分丝状藻类、水草碎屑等。2龄鱼可达性成熟,产卵时期在5-6月,生殖时常成群逆水而上,到达底质为砂和砾石的急流处产卵。卵呈黏性,产出的卵粘附在岩石或水草上发育。江河、湖泊、水库中皆有分布(图4-26)。

细鳞鲴(Xenocypris microlepis)
生活习性:喜生活于水的中下层,生殖时期常集群溯水而上,到通湖(库)的河流急流处进行生殖。性成熟年龄为2龄,亦有3龄鱼才达性成熟的(体长为305-390 mm、体重为413-1100 g)。生殖时期在 5-8 月,多集中在 5-6 月。卵粘性,分批产出。成鱼以硅藻、丝状藻以及植物碎屑为食;幼鱼多以浮游生物为食。分布在长江和汉水干支流、水库、湖泊(图4-27)。

(3)肉鱼性鱼类增殖放流
肉食性鱼类能够摄食水体中的小鱼小虾和底栖动物进行能量补充,其强大的捕食作用能通过下行效应对湖泊生态系统的其他生物造成影响。肉食性鱼类摄食小型鱼类,会导致浮游动物在一段时间大量繁殖,进而造成浮游植物数量急剧减小,进而能改改善水质。同时浮游植物的减少能够降低水体发生水华的几率,减少水体承载的负荷,促进湖泊生态的恢复。重点增殖放流以鳜为主的包括翘嘴鮊在内的凶猛食鱼性鱼类,通过摄食小鱼小虾,减少能量的外泄的同时还能够增长自身的经济价值。黄颡鱼作为底栖动物食性的鱼类,也可以辅助进行增殖放流,合理利用湖泊的底栖生物资源。基于西凉湖供饵能力及同类湖泊的经验,确定的生态放养方案如下:
放养时间:夏花寸片鱼种一般在6月至7月初,冬片鱼种一般在12月至翌年2月初。
放养规格和密度:搭配不同种类的食鱼性鱼类能够充分利用水体空间,为了提高经济效益,拟定投放鳜、翘嘴鮊、黄颡鱼等物种,结合西凉湖目前所含有的种质资源状况,建议2021-2025年放养规格、数量如下(表4-13)。
鳜(Siniperca chuatsi)
生活习性:一般喜栖息在静水水域,尤以水草丛生处较多,冬季喜在深水处越冬。许多湖区渔民,常依据鳜鱼喜侧卧水底凹陷处的习性,采用“踩鳜鱼”的渔法,可大量捕获。鳜为凶猛性鱼类,主要以捕食其它小鱼类为主,亦食部分水生昆虫、虾类。雌鱼一般2龄鱼可达性成熟,生殖时期5-7月,卵浮性。分布:鳜的分布较广,各河流干流及支流、湖泊、水库中常见(图4-28)。

翘嘴鲌(Culter alburnus)
生活习性:喜生活于水中的中上层,游动迅速,善跃。成鱼完全以小型鱼类为食,为凶猛性鱼类。2-3龄可达到性成熟,一般6月中旬开始产卵,卵稍呈粘性。分布在长江、汉水,以及湖泊、水库中(图4-29)。

蒙古鲌(Culter mongolicus)
生活习性:喜生活于水的中上层。幼鱼时以摄食小型动物,如水生昆虫、大型浮游生物等为食,随个体增大,则以小鱼为主食,常见食物鱼有䱗条、鲫、似鳊等。2龄鱼可达性成熟。生殖时期多集中在5月底至7月。卵粘性。分布:长江干流及汉水,湖泊和水库皆有(图4-30)。

黄颡鱼(Pelteobagrus fulvidraco)
生活习性:喜生活在水体的底层,鱼苗阶段以浮游动物为食,成鱼则以昆虫及其幼虫、小鱼虾、螺蚌等为食。一般至1龄大部分性成熟,2龄全部性成熟,3龄以后体长相对增长率递减明显。分布于珠江、闽江、湘江、长江、黄河(图4-31)。

物种类型 | 放养时间 | 放养规格 | 投放地点 | |
滤食性鱼类 | 鲢 | 鱼种:1-2月份 夏花:6-7月份 | 大规格:0.5 kg/尾 中规格:0.2 kg/尾 寸片:约100尾/kg | 湖心 |
鳙 | ||||
肉食性鱼类 | 鳜 | 夏花寸片:6-7月初 冬片:12月至翌年2月初 | 10-26 g/尾 | 远离进出水口;水生植物较丰富的沿岸区 |
翘嘴鲌 | 远离进出水口,生境异质性高、饵料资源丰富的水域 | |||
蒙古鲌 | 远离进出水口;水生植物较丰富区域 | |||
黄颡鱼 | 远离进出水口,饵料资源丰富的水域 | |||
碎屑食性鱼类 | 细鳞鲴 | 夏花:6月份 冬片:冬季捕捞结束或次年3月份前 | 夏花:3-4 cm 冬片:10-25 g | 远离进出水口,宜选择生境异质性高、饵料资源丰富的水域 |
黄尾鲴 | ||||
时间 | 种类 | 放养规格 | 放养数量 |
2021年 | 鲢(大规格) | 400-500 g | 280万斤 |
鳙(大规格) | 400-500 g | 140万斤 | |
鳜 | 10-25 g | 8万尾 | |
细鳞鲴 | 10-25 g | 10万尾 | |
黄尾鲴 | 10-25 g | 10万尾 | |
翘嘴鲌 | 10-25 g | 8万尾 | |
蒙古鲌 | 10-25 g | 8万尾 | |
黄颡鱼 | 10-25 g | 10万尾 | |
2022年 | 鲢(大规格) | 400-500 g | 150万斤 |
鳙(大规格) | 400-500 g | 80万斤 | |
鲢(中规格) | 200-250 g | 80万斤 | |
鳙(中规格) | 200-250 g | 40万斤 | |
细鳞鲴 | 10-25 g | 6万尾 | |
蒙古鲌 | 10-25 g | 6万尾 | |
鳜 | 10-25 g | 4万尾 | |
黄颡鱼 | 10-25 g | 5万尾 | |
2023年 | 鲢(大规格) | 400-500 g | 150万斤 |
鳙(大规格) | 400-500 g | 80万斤 | |
鲢(寸片) | 100尾/kg | 150万尾 | |
鳙(寸片) | 100尾/kg | 80万尾 | |
黄尾鲴 | 10-25 g | 6万尾 | |
翘嘴鲌 | 10-25 g | 6万尾 | |
黄颡鱼 | 10-25 g | 5万尾 | |
鳜 | 10-25 g | 4万尾 | |
2024年 | 鲢(大规格) | 400-500 g | 150万斤 |
鳙(大规格) | 400-500 g | 80万斤 | |
鲢(中规格) | 200-250 g | 80万斤 | |
鳙(中规格) | 200-250 g | 40万斤 | |
细鳞鲴 | 10-25 g | 6万尾 | |
蒙古鲌 | 10-25 g | 6万尾 | |
黄颡鱼 | 10-25 g | 5万尾 | |
鳜 | 10-25 g | 4万尾 | |
2025年 | 鲢(大规格) | 400-500 g | 150万斤 |
鳙(大规格) | 400-500 g | 80万斤 | |
鲢(寸片) | 100尾/kg | 150万尾 | |
鳙(寸片) | 100尾/kg | 80万尾 | |
黄尾鲴 | 15-30 g | 6万尾 | |
翘嘴鲌 | 4-6 cm | 6万尾 | |
黄颡鱼 | 8-10 cm | 5万尾 | |
鳜 | 8-10 cm | 4万尾 |
4.2.9.2 鱼类合理捕捞方案
鱼类合理捕捞能够调节鱼类群落结构,定期捕捞主要遵循以下两点:一是合理的捕捞能够充分利用湖泊资源,捕捞生长年限较长的鱼类资源,并补充一批优质的鱼苗,能够提高生态系统的稳定。二是合理科学的捕捞能够使鱼群处在生长的最大趋势,既能提高社会经济效益,又能保持湖泊中鱼类群落不遭到破坏。
针对西凉湖相关政策,捕捞作业应该遵循相关规定。根据不同食性的鱼类制定不同的捕捞方案(表4-14):
滤食性鱼类捕捞:一般在当年的9月份开始,大规模捕捞宜在11月下旬至翌年1月。捕捞主要有联合渔法,也可采用刺网捕捞,但应严格控制网目大小,做到留小捕大。建议鲢鱼最小捕捞个体规格大于1 kg,鳙鱼最小捕捞个体规格为1.5 kg。
碎屑食性鱼类捕捞:首次捕捞在放养后第二年9月份开始;以后每年4月-5月可进行小规模捕捞,10月至翌年1月可进行大规模捕捞。 捕捞主要有联合渔法,也可采用刺网和网簖捕捞,但应严格控制网目大小,最小捕捞个体规格为0.5 kg。
肉鱼性鱼类捕捞:首次捕捞在放养后第二年9月份开始;以后每年4-5月可进行小规模捕捞,10月至翌年1月可进行大规模捕捞。捕捞主要有联合渔法,也可采用刺网和网簖捕捞,但应严格控制网目大小。鳜最小捕捞个体规格为0.5公斤,翘嘴鲌和蒙古鲌为0.5公斤,黄颡鱼为0.15公斤。
时间 | 捕捞种类 | 捕捞季节 | 最小起捕规格 | 最大年捕捞量 |
2021 | 鲢 | 11月-1月 | 1 kg/尾 | 840万斤 |
鳙 | 11月-1月 | 1.5 kg/尾 | 420万斤 | |
鳜 | 4月-5月、10月-1月 | 0.5 kg/尾 | 2万斤 | |
细鳞鲴 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
黄尾鲴 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
翘嘴鲌 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
蒙古鲌 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
黄颡鱼 | 4月-5月、10月-1月 | 0.15 kg/尾 | 5万斤 | |
2022 | 鲢 | 11月-1月 | 1 kg/尾 | 650万斤 |
鳙 | 11月-1月 | 1.5 kg/尾 | 360万斤 | |
鳜 | 4月-5月、10月-1月 | 0.5 kg/尾 | 2万斤 | |
细鳞鲴 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
蒙古鲌 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
黄颡鱼 | 4月-5月、10月-1月 | 0.15 kg/尾 | 5万斤 | |
2023 | 鲢 | 11月-1月 | 1 kg/尾 | 600万斤 |
鳙 | 11月-1月 | 1.5 kg/尾 | 300万斤 | |
黄尾鲴 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
鳜 | 4月-5月、10月-1月 | 0.5 kg/尾 | 2万斤 | |
翘嘴鲌 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
黄颡鱼 | 4月-5月、10月-1月 | 0.15 kg/尾 | 5万斤 | |
2024 | 鲢 | 11月-1月 | 1 kg/尾 | 600万斤 |
鳙 | 11月-1月 | 1.5 kg/尾 | 300万斤 | |
细鳞鲴 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
蒙古鲌 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
黄颡鱼 | 4月-5月、10月-1月 | 0.15 kg/尾 | 5万斤 | |
鳜 | 4月-5月、10月-1月 | 0.5 kg/尾 | 2万斤 | |
2025 | 鲢 | 11月-1月 | 1 kg/尾 | 600万斤 |
鳙 | 11月-1月 | 1.5 kg/尾 | 300万斤 | |
黄尾鲴 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
翘嘴鲌 | 4月-5月、10月-1月 | 0.5 kg/尾 | 4万斤 | |
黄颡鱼 | 4月-5月、10月-1月 | 0.15 kg/尾 | 5万斤 | |
鳜 | 4月-5月、10月-1月 | 0.5 kg/尾 | 2万斤 |
4.2.10.1 藻类水华形成与危害
水华是淡水中的一种自然生态现象。绝大多数的水华是仅由藻类引起的如蓝藻、绿藻、硅藻等也有部分的水华现象是由浮游动物—腰鞭毛虫引起的。水华发生是多种因素共同作用的结果。这些因素包括 :①水体中丰富的N 、P营养物质;②适宜的温度和光照 ;③适宜的水文地理条件如缓慢的水流等,但是最主要的原因还是氮磷等营养元素超标。2018-2019年,西凉湖西湖区总氮、总磷含量超标,具有潜在发生水华的危险。水华形成的危害最主要的是造成水体缺氧,大量藻类增殖消耗水体中的氧气,并且覆盖在水体表面,使水体以下难以接触到阳光,对水生生物的生长发育不利;甚至像微囊藻发生水华时,还能产生毒素,毒害水生生物资源,并且也会危害人体的健康。
4.2.10.2 藻类水华预防措施
对于藻类水华爆发之前,我们要时刻关注水的健康,具体有以下几点:
1.强化监测预警。严密监控重点湖库水质和蓝藻水华动态,在蓝藻水华暴发敏感区和高发期,制定和实施加密监测方案,综合水文、气象、水质和藻类监测结果,加强蓝藻水华形势分析和预测预警。
2.制定应急预案。对发生过规模化蓝藻水华的重点湖库,应制定蓝藻水华防控应急预案,建立应急工作机制,明确责任分工,加强应急物资储备,定期开展应急预案演练。
3.加强敏感水域水华控制。对于集中水源地取水口及其周边至少500米范围区域,开展蓝藻围挡和打捞处置。加强饮用水水源水质监控,及时分析蓝藻水华对饮用水安全的影响。
4.实施水华综合防控。持续推进重点湖库生态环境治理,对汇入富营养化湖库的河流应严格实施氮磷浓度控制,及时清理入湖河沟淤泥、垃圾,降低入湖污染负荷。
4.2.10.3 藻类水华的治理措施
藻类水华的治理措施主要有生物、化学和物理
1.物理方法主要通过人工打捞的方法,将湖泊水库中的藻类清除干净,但是此项工作任务量大,并且不容易从源头解决问题。
2.化学方法主要是通过化学试剂等杀死水体中的藻类,此方见效快,但是极易污染水体,给水体第二次加重负荷。
3.生物方法主要是通过生物调控,在水体中通过放养滤食性鱼 类,通过鱼类的摄食作用,减轻水华带来的危害作用。
本次规划设计了十大生态修复工程:1)湖区周围环境治理工程、2)沉水植物修复工程、3)挺水植物修复工程、4)浮叶植物修复工程、5)立体生态平台建设工程、6)水葫芦治理工程、7)软体动物修复工程、8)关键栖息地修复工程、9)鱼类群落调控工程、10)藻类水华预防与处理工程。上述工程的实施将优化西凉湖水生生物群落结构、提高水域生态系统多样性,对改善湖区水质,促进湖区生态经济效益的发展有积极作用
根据所设计工程项目的具体内容工程投资预算等内容,形成了西凉湖生态修复规划项目库,见表5-1。
序号 | 项目名称 | 建设内容 | 投资(万元) |
1 | 湖区周围环境治理工程 | 1)控制农业面源污染,严格规范化肥使用;2)清理湖区垃圾,杜绝生活垃圾入湖;3)控制污染物排放,建立污染物排放监控和处理机制。 | 4000 |
2 | 沉水植物修复工程 | 建立沉水植物修复区:苦草4.72 km2,穗花狐尾藻1.42 km2,轮叶黑藻2.83 km2,金鱼藻0.94 km2,马来眼子菜0.10 km2。 | 1800 |
3 | 挺水植物修复工程 | 建立挺水植物修复区:狄叶香蒲4.67 km2,芦苇3.00 km2,莲2.00 km2,水葱0.33 km2。 | 1000 |
4 | 浮叶植物修复工程 | 建立浮叶植物修复区:荇菜4.29 km2,水鳖1.43 km2,芡实0.29 km2。 | 800 |
5 | 立体生态平台建设工程 | 在西凉湖污染较为严重的区域设置立体生态平台,在去除污染物同时能够达到净水的效果,维持水环境健康。 | 300 |
6 | 水葫芦治理工程 | 建立面积为8 km2的水葫芦治理区域,将水葫芦集中后进行集中围栏处理,通过放养草鱼和打捞等综合手段进行治理。 | 1000 |
7 | 软体动物修复工程 | 建立软体动物修复区:螺类6.6 km2,蚌类6.6 km2 | 800 |
8 | 关键栖息地修复工程 | 使用人工鱼巢在湖泊中建立人工产卵场共计8 hm2,修复鱼类栖息地,辅助鱼类产卵。 | 300 |
9 | 鱼类群落调控工程 | 通过进行鱼类增殖放流,对水质和水生生物群落进行调控,以2021年为例,投放滤食性鱼类420万斤、碎屑食性鱼类20万尾、底层食鱼性鱼类18万尾、上层食鱼性鱼类16万尾。 | 4500 |
10 | 藻类水华预防与处理工程 | 在藻类水华爆发前,通过监测、制定预案、优化生物群落结构、加强环境治理等措施进行预防;藻类水华发生时积极采用各种手段进行之类。 | 2500 |
投资总计(万元) | 17000 | ||
十九大报告以来,生态文明已逐渐受到国家的重视,国家、政府部门以及地方管理部门对生态环境越来越予以高度重视,本次规划合理调整了湖泊鱼类群落的结构、提高了水生植物的多样性,通过生物操作等手段,恢复大型水生植物、沉水植物以及种质资源量,为保护水资源,恢复生态环境健康提供良好的条件。本次规划的实施,有利于恢复西凉湖生态环境,促进西凉湖水生物资源的可持续性,维持西凉湖生态文明具有重大意义。
6.1.1.1 移除氮磷等营养物质,优化水域生态环境
鱼类是生态系统中重要的组成成分,是水体中的消费者,不同类群的鱼群既可以摄食浮游生物、底栖生物、水生植物,还可以摄食水体中碎屑残渣进行调控水体。按照食物链食物网的原理,合理分配利用水中生物资源,通过以生物技术为主,同时配合物理技术和化学技术,达到实现生态修复的最大效益。在实施生态修复的同时,合理的调整鱼类群落结构,在放养鲢、鳙等滤食性鱼类的同时,搭配食鱼性(鳜、翘嘴鮊等)共同净化水体,移除水体中的氮磷,草食性(草鱼等)控制水葫芦等有害植物的生物量,恢复水体健康,在保证生态修复的同时,有效的提高渔业效率。
当前湖泊中,可以直接利用水体中游离氮磷元素的生物类群主要为浮游生物,而浮游生物食性的鲢和鳙为主要增殖放流对象,鲢、鳙可通过大量摄食浮游生物,从而移除水体中的氮磷元素,降低湖泊富营养化水平。通过研究鲢、鳙鱼体中氮、磷元素的含量,可以计算出鲢每增重1 g,可以从水体中移除23.2 mg的氮和5.8 mg的磷;鳙每增重1 g可以从水体中移除18 mg的氮和7.7 mg的磷。根据西凉湖鱼类放养的情况,经过一年的生长,预计规格为500 g的鲢、鳙苗体重可实现2-3倍的增长。因此,按照本规划的实施目标,鲢、鳙经过一周年的投放和生长,其鱼体增重和移除水体氮、磷含量见表6-1。假设以最低增长重量计算(即增重2倍),按2021年西凉湖鲢、鳙放养量,可分别实现240万kg和140万kg的增重,2021年西凉湖鲢和鳙可贮存氮80.88 t、磷24.70 t,相当于西凉湖全湖水体氮含量可下降0.46 mg/L,磷含量可下降0.14 mg/L。
种类 | 放养量(万斤) | 增重(万kg) | 每克移 除氮(mg) | 每克移 除磷(mg) | 共移 除氮( t) | 共移 除磷( t) |
鲢 | 240 | 240 | 23.2 | 5.8 | 55.68 | 13.92 |
鳙 | 140 | 140 | 18 | 7.7 | 25.20 | 10.78 |
总计 | 80.88 | 24.70 | ||||
6.1.1.2 优化鱼类群落结构,提高生物多样性
水生态修复与保护主要以生物操纵的方法,通过上行和下行效应作用,对水体中浮游生物加以控制,达到改善水体的作用。提高鱼类群落的多样性有利于丰富食物链、食物网结构,适当的搭配名特优品种如鳜、黄颡鱼等,能够充分利用水体资源,并在生态修复与保护过程中达到生态-经济效益双丰收。
6.1.1.3恢复湖泊生态系统结构,提高水生生物多样性
通过对湖泊生态修复与保护,有效的保护水资源,并全面恢复浮游植物、浮游动物、底栖动物、水生高等植物和鱼类群落,优化水体生态系统结构,促进物质循环和能量转换。本项目不仅恢复了水生物资源,而且通过水生高等植物种植方法,对水体中沉水植物、浮叶植物进行修复,大大提高了湖区水生高等植物的覆盖率。水生高等植物的修复在一定程度上能够对食浮游生物的小型鱼类加以控制,进而能够提高浮游植物和浮游动物的多样性。
西凉湖水生态修复与保护的实施,不仅保护了西凉湖水生物资源,而且有利于周围居民的发展。大型水生植物具有景观效果,能够带动当地旅游产业的发展,鱼类资源的增殖能够吸引更多爱垂钓的人士,推动了当地经济的发展。水生态环境变好能够改善附近居民的生活环境,此外,还能够吸引更多的人前来观赏,同时也提高了当地餐饮行业的发展。
市场风险往往是由供求关系和价格变化所构成,每年养殖数量和养殖品种的变化,都会影响到市场价格的波动,但随着近些年来,养殖模式以及养殖技术的不断成熟,在一定程度上能够保证每年水产品进入市场的规模,减轻了由于价格波动带来的市场风险。
自然风险主要是指天气气候等原因给渔业生产造成损失,自然界频繁发生的自然灾害,常常给渔业生产带来重大损失。随着现代科技的发展,自然灾害能够通过科技力量提前预测,并采取相关措施减小损失。本规划在预防灾害方面做了充分的考虑,强调提高管理效率以减轻人类活动造成的不利影响。加强各种管控措施以期将自然风险将至最低。
技术风险主要由于在操作过程中出现失误甚至出错,导致技术风险的增加,针对技术风险,可以通过培训,增加技术人员对相关操作更加熟练,在减少技术风险的同时还能提高生产效益。
管理风险一般是由于管理者或生产者计划、组织和控制不当或失误给渔业生产造成损失的可能性,管理风险可通过完善渔业管理制度,要求管理人员加强管理力度,选择合适的管理方式,保证在管理工作中不出现任何疏漏。
湖泊水生态修复与保护需要贯彻在不破坏自然环境的前提下,利用生物资源对水体进行修复与保护,坚持人和自然和谐发展,对区域公共管理进行深化改革、创新体制;坚持水生态修复对水生物资源的可持续发展。高度重视建设规划实施方案的制定和修订工作,拟定实施措施。实施过程中严格按规划建设方案,认真组织实施,确保方案按既定要求如期完成。
充分认识到湖泊水生态修复与保护的战略地位以及生态环境对区域经济发展、百姓就业致富等方面的巨大作用,进一步提高全社会重视渔业水域和鱼类种质资源保护及开发利用意识,切实加强对项目建设的领导,相关措施有以下两点:
1.政府各部门与当地执法机构进行协调,制定严格的工作机制,将西凉湖水生态修复纳入目标考核内容当中,对执法过程中违反相关规定的人员予以不同程度的处罚。
2.严格按照相关规定进行执法,做到严格公正、不疏漏,形成齐抓共管局面。
建立以国家和地方政府资金投入为导向,根据西凉湖生态修复后所带来的经济效益,积极倡导企业和生产者投入为主体,金融信贷为支撑等方式,广辟资金渠道,加大招商引资,强化资金投入与管理。同时鼓励广大群众响应生态修复与保护规划工程,以水面股份制等入股方式为西凉湖生态发展提供充足的资金保障。
根据湖区在进行水生态修复与保护过程的技术薄弱环节,一是注重成熟生产技术的学习,通过与科研院所和高校紧密联系,逐步提高相关技术人员专业结构,优化和提高技术人员的专业知识。二是借鉴先进技术资料,取长补短,将先进的科研技术成果应用到实际当中。三是积极鼓励科技创新,将更高效的水生态修复技术应用到湖泊治理当中,促进湖区水生态修复与保护发展。
完善环境与工程施工综合决策机制,强化环境综合整治目标管理责任制,建立统一监督管理机制和反馈机制,完善违法行为责任追究制度,加强渔业执法体系建设,健全统一指挥、责权明确、行为规范、监督有效、保障有力的渔业行政执法体制,进一步完善配套渔业执法设施、设备,加大渔业行政执法力度,全面实施水产种苗生产经营许可制度、水产品检疫制度、水产品市场管理制度、“禁渔”和捕捞许可制度等,保障湖区水生态修复与保护的发展。
 鄂公网安备 42120202000229号 网站地图
鄂公网安备 42120202000229号 网站地图

CopyRight © 2018 咸宁市农业农村局
技术支持:咸宁新闻网